Bảng 3.10: Tổng hợp kết quả giõm hom các cây trội Keo tai tượng
Cây trội | Sè hom giâm | Sè hom ra rÔ | Tỷ lệ ra rễ trung bình (%) | V% | Vị trí cắt | |
1 | K4 | 110 | 40 | 36,1 | 25,1 | 1 (1,2m) |
2 | K101 | 992 | 806 | 84,2 | 8,5 | 3 (Cắt cành dưới tán) |
3 | K102 | 863 | 680 | 81,1 | 7,4 | |
4 | K112 | 344 | 232 | 68,2 | 3,4 | |
5 | K77 | 228 | 150 | 66,4 | 5,5 | |
6 | K113 | 147 | 94 | 64,2 | 2,2 | |
7 | K88 | 231 | 145 | 62,1 | 5,6 | |
8 | K107 | 121 | 74 | 61,4 | 7,0 | |
9 | K87 | 103 | 61 | 59,2 | 3,1 | |
10 | K75 | 245 | 132 | 55,2 | 4,6 | |
11 | K97 | 464 | 238 | 52,0 | 5,0 | |
12 | K93 | 392 | 197 | 50,7 | 3,1 | |
13 | K99 | 162 | 77 | 48,1 | 8,0 | |
14 | K73 | 155 | 69 | 45,0 | 6,5 | |
15 | K100 | 285 | 122 | 43,3 | 7,5 | |
16 | K116 | 453 | 174 | 39,0 | 5,8 | |
17 | K84 | 209 | 73 | 35,6 | 7,0 | |
18 | K111 | 826 | 118 | 14,6 | 6,2 | |
Céng | 6.220 | 3.442 | ||||
19 | K98 | 170 | 100 | 59,7 | 5,6 | 4 (Cắt cụt ngọn) |
20 | K83 | 378 | 201 | 54,1 | 8,2 | |
21 | K82 | 521 | 263 | 51,1 | 3,5 | |
22 | K85 | 524 | 238 | 45,5 | 7,0 | |
23 | K90 | 821 | 343 | 41,4 | 5,5 | |
24 | K94 | 179 | 69 | 38,7 | 8,0 | |
25 | K80 | 359 | 136 | 37,9 | 3,6 | |
26 | K86 | 254 | 87 | 34,1 | 4,9 | |
27 | K72 | 231 | 79 | 33,9 | 5,3 | |
28 | K92 | 753 | 246 | 32,1 | 6,5 | |
29 | K79 | 232 | 73 | 31,3 | 5,6 | |
30 | K78 | 191 | 55 | 29,2 | 4,5 | |
31 | K109 | 552 | 128 | 23,0 | 2,8 | |
32 | K81 | 471 | 106 | 22,7 | 4,8 | |
33 | K91 | 330 | 74 | 22,6 | 4,6 | |
34 | K104 | 250 | 52 | 20,9 | 3,1 | |
35 | K89 | 91 | 17 | 18,7 | 9,5 | |
36 | K96 | 115 | 19 | 16,6 | 1,2 | |
37 | K110 | 272 | 43 | 15,8 | 5,3 | |
Céng | 6.694 | 2.329 |
Có thể bạn quan tâm!
-

 Phương Pháp Phân Tích Một Số Tính Chất Cơ Lý Gỗ
Phương Pháp Phân Tích Một Số Tính Chất Cơ Lý Gỗ -
 Độ Co Rút, Độ Hút Nước & Dãn Dài, Độ Hút Ẩm, Độ Bền Nén Dọc Và Độ Bền Uốn Tĩnh
Độ Co Rút, Độ Hút Nước & Dãn Dài, Độ Hút Ẩm, Độ Bền Nén Dọc Và Độ Bền Uốn Tĩnh -
 So Sánh Tính Đồng Nhất Về Các Tính Chất Cơ Lý Gỗ
So Sánh Tính Đồng Nhất Về Các Tính Chất Cơ Lý Gỗ -
 Chọn cây trội, dẫn dòng Keo tai tượng Acacia mangium Wild và ứng dụng công nghệ sinh học trong bố trí thí nghiệm và xây dựng vườn giống - 8
Chọn cây trội, dẫn dòng Keo tai tượng Acacia mangium Wild và ứng dụng công nghệ sinh học trong bố trí thí nghiệm và xây dựng vườn giống - 8
Xem toàn bộ 69 trang tài liệu này.
Ngoài ra, với một số cây có điều kiện cắt ở cả 2 vị trí, đề tài đã thí nghiệm giâm hom cho 2 loại hom của một cây: hom có nguồn gốc từ cành “C” (vị trí 3) và hom ngọn “N” (vị trí 4). Kết quả được tổng hợp ở bảng sau:
![]()
![]()
![]()
![]()
Bảng 3.11: Tổng hợp kết quả giâm hom theo vị trí lấy hom
Cây trội | Số hom giâm | Sè hom ra rÔ | Tỷ lệ ra rễ trung bình (%) | V% | |
1 | K72 N K72 C | 231 105 | 79 63 | 33.9 59.9 | 5.3 4.5 |
2 | K78 N K78 C | 191 114 | 55 66 | 29.2 57.9 | 4.5 2.3 |
3 | K80 N K80 C | 359 149 | 136 94 | 37.9 63.5 | 3.6 3.4 |
4 | K81 N K81 C | 471 99 | 106 49 | 22.7 49.6 | 4.8 3.6 |
5 | K83 N K83 C | 378 117 | 201 94 | 54.1 80.5 | 8.2 1.6 |
6 | K86 N K86 C | 254 113 | 87 72 | 34.1 63.9 | 4.9 2.7 |
7 | K90 N K90 C | 821 135 | 343 96 | 41.4 71.5 | 5.5 7.0 |
8 | K98 N K98 C | 170 105 | 100 86 | 59.7 81.9 | 5.6 0.9 |
![]()
![]()
![]()
![]()
Kết quả tổng hợp ở bảng 3.11 cho thấy tất cả các hom có nguồn gốc từ các cành phía dưới (vị trí 3) đều có tỷ lệ ra rễ cao hơn rất nhiều so với các hom có nguồn gốc từ ngọn (vị trí 4). Đặc biệt là dòng K83 C và K98 C có tỷ lệ ra rễ rất cao (>80%). Điều này một lần nữa chứng minh lý thuyết về đặc điểm đặc trưng là “Hom càng xa gốc tuổi sinh lý càng già thì hom càng khó ra rễ”.
Nhìn chung Keo tai tượng là đối tượng rất khó ra rễ, đặc biệt là các cây
đã đạt tuổi thành thục. Nhưng theo yêu cầu của công tác chọn giống cây rừng, rừng để chọn lọc cây trội phải ở tuổi gần thành thục hoặc thành thục công nghệ, ở tuổi này các cây trội mới thể hiện đầy đủ các đặc điểm ưu việt. Do vậy
việc dẫn dòng và giâm hom thành công cho các cây trội Keo tai tượng 10 - 13 tuổi có ý nghĩa rất lớn trong công tác chọn giống Keo tai tượng.
3.2. Kết quả phân tích đa dạng di truyền của các cây trội đã chọn và ứng dụng kết quả này trong bố trí thí nghiệm và xây dựng vườn giống
Trong một thời gian dài, di truyền phân tử đã từng được coi là một lĩnh vực khoa học mang tính lý thuyết thuần tuý nhưng nay đã trở thành một công cụ ứng dụng thực tế rộng rãi, nòng cốt của cuộc cách mạng công nghệ sinh học đang diễn ra trên phạm vi toàn cầu. Có được như vậy là nhờ những phát minh đột phá trong lịch sử phát triển của Di truyền phân tử nửa cuối thế kỷ 20. Trên cơ sở những thành tựu đó đã ra đời hàng loạt những kỹ thuật thao tác ADN. Trong đó phản ứng chuỗi trùng hợp PCR đã trở nên phổ biến và hữu dụng trong nghiên cứu và trong sản xuất.
Kỹ thuật nhân ADN đặc hiệu, còn gọi là chuỗi phản ứng trùng hợp hay kỹ thuật PCR được Kary Mullis hoàn thiện vào giữa những năm 80 và đã đem
đến sự tiến bộ ở tất cả các lĩnh vực của sinh học hiện đại, trong đó chỉ thị phân tử đóng vai trò ngày càng quan trọng trong nghiên cứu đa dạng di truyền, quan hệ tiến hoá giữa các loài, xác định các loài và giống [44].
Trong nghiên cứu đa dạng di truyền, chỉ thị RAPD (Đa hình các đoạn ADN nhân ngẫu nhiên) được dùng phổ biến vì kỹ thuật này đơn giản, ít tốn kém. Còn để nhận dạng loài hay xuất xứ thường dùng chỉ thị lục lạp là do lục lạp có tính bảo thủ cao và quan trọng hơn nữa là tần số đột biến thấp hơn so với ADN nhân, do đó sự đa hình của ADN lục lạp (cpADN) được sử dụng trong nghiên cứu về hình thái và quan hệ loài như ở các chi Cistrus, Hemerocallis, Gordonia và Polyspora [41].
Trong khuôn khổ luận văn này, đề tài xin trình bày một số kết quả phân tích chỉ thị phân tử và đánh giá quan hệ di truyền giữa các dòng Keo tai tượng từ đó lựa chọn cặp bố mẹ trong bố trí thí nghiệm vườn giống.
3.2.1. Kết quả phân tích với các mồi RAPD
Sản phẩm PCR với các mồi RAPD được điện di trên gel agarose 0,8%, kết quả thể hiện trong các hình sau:
Hình 3.10: Sản phẩm PCR với mồi OPB17 với ADN genome của một số dòng Keo tai tượng
Hình 3.11: Sản phẩm PCR với mồi OPC18 với ADN genome của một số dòng Keo tai tượng
Các sản phẩm PCR ADN genome của các dòng Keo tai tượng với các mồi RAPD cho thấy tất cả các mồi đều cho đa hình nhưng mức độ đa hình thấp. Trong 5 mồi sử dụng có các mồi OPC18 và OPB17 cho nhiều băng đa hình hơn (hình 3.10 và 3.11)
3.2.2. Kết quả phân tích với mồi lục lạp trnL
Hình 3.12: Sản phẩm PCR mồi lục lạp trnL với ADN genome của một số dòng Keo tai tượng
M A7 A31 C W P I K4 K72 K75 K77 K78 K79 K80 K81
M K82 K83 K84 K85 K86 K88 K89 K90 K91 K92 K93 K94 K96 K97
1000 bp -
500 bp -
400 bp -
300 bp -
200 bp -
Hình 3.13: Sản phẩm PCR mồi lục lạp trnL với ADN genome của một số dòng Keo tai tượng cắt bằng TaqI
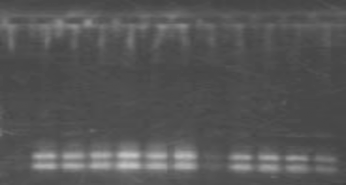
K98 K100 K101 K102 K104 K107 K109 K110 K111 K112 K113 K116
Hình 3.14: Sản phẩm PCR mồi lục lạp trnL với ADN genome của một số dòng Keo tai tượng cắt bằng TaqI
Trên hình 3.12 ta thấy về độ dài của sản phẩm PCR của các dòng Keo tai tượng với mồi trnL không có đa hình. Điều này cho thấy mối quan hệ di truyền rất gần nhau giữa các dòng Keo tai tượng. Để có thể phân tích sâu hơn về sự khác nhau giữa các dòng Keo tai tượng, sản phẩm PCR với mồi trnL
được cắt với enzym TaqI với mục đích tìm ra sự khác nhau trong trình tự ADN, kết quả thể hiện trong hình 3.13 và 3.14. Sau khi cắt bằng enzym TaqI sản phẩm PCR của các dòng Keo tai tượng đã xuất hiện đa hình. Điều này
chứng tỏ một số dòng Keo tai tượng đã có sự khác nhau trong trình tự gen trnL. Ngoài ra với gen trnL có thể phân biệt được xuất xứ Carwell (C1) nhờ không có phân đoạn 700bp (hình 3.13 và 3.14). Các dòng không có phân đoạn 700bp là K79, K80, K81, K83, K86, K88, K90, K91, K92, K93, K96, K97 và K100. Qua
phân tích trên cho thấy 13 dòng này thuộc xuất xứ Carwell và được trồng vào
năm 1993 ở Lâm trường Nguyễn Văn Trỗi thuộc tỉnh Tuyên Quang.
3.2.3. Quan hệ di truyền giữa các dòng Keo tai tượng và lựa chọn cặp bố mẹ trong xây dựng vườn giống
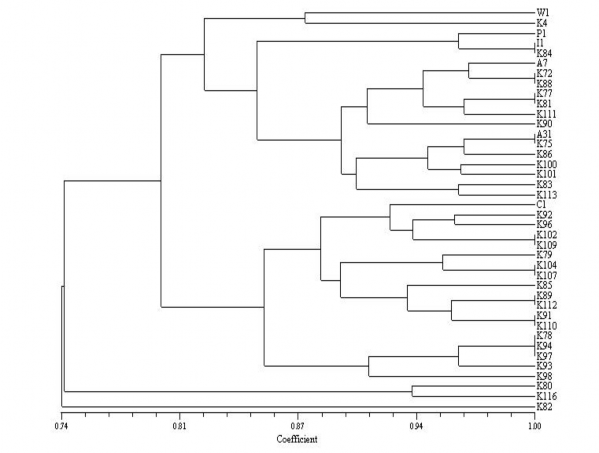
Quan hệ di truyền giữa các dòng Keo tai tượng được thể hiện như sau:
Hình 3.15: Biểu đồ quan hệ di truyền giữa các dòng Keo tai tượng
Qua biểu đồ quan hệ di truyền trên ta cũng thấy các dòng keo có mối quan hệ di truyền rất gần nhau (hệ số tương đồng di truyền lớn hơn 70%). Các
kết quả phân tích này cũng phù hợp với ước đoán di truyền trên cơ sở isoenzyme ở các loài keo của Monran và cộng sự 1988 [43], theo ông hệ số di truyền (he) của Keo tai tương là rất thấp chỉ đạt 0,017 so giá trị trung bình của loài keo là 0,147. Sở dĩ tại nơi nguyên sản Keo tai tượng vẫn có đa dạng di truyền hẹp là do chúng có phạm vi phân bố và phân bố không liên tục, chúng thường gặp trong các quần thể rất nhỏ [47].
Nhìn vào hình 3.15 có thể chia các dòng Keo tai tượng làm 2 nhóm chính có quan hệ di truyền gần nhau ở mức gần 80%:
+ Nhóm I gồm 5 xuất xứ W1, P1, I1, A7, A31 và các dòng K4, K84, K72, K88, K77, K81, K111, K90, K75, K86, K100, K101, K83, K113.
+ Nhóm II gồm 1 xuất xứ C1 và các dòng K92, K96, K102, K109, K79, K104, K107, K85, K89, K112, K91, K110, K78, K94, K97, K93, K98.
Riêng 3 dòng K80, K116 và K82 có quan hệ di truyền xa hơn cả so với tất cả các dòng khác với mức tương đồng từ 17 - 40%. Về các xuất xứ: Xuất xứ P1 và I1 gần nhau nhất (giống nhau tới trên 95%) tiếp đến là A7, A31 (giống nhau 89%) và W1 (gièng P1, I1, A7, A31 đến 80%). Riêng xuất xứ C1 nằm khác nhóm, mức độ khác biệt từ 16 - 28% so với các xuất xứ khác.
Nhìn vào hình 3.15 ta có thể lựa chọn các cặp bố mẹ có khoảng cách di truyền xa nhau để trồng sát nhau. Tránh trồng những cặp có khoảng cách di truyền giống nhau như I1 với K84, K72 với K88, K77 với K81, K75 với Am31, K102 với K109... cạnh nhau nhằm trách thụ phấn cận huyết hay nói một cách khác là làm tăng thu di truyền ở thế hệ sau; nhất là đối với loài Keo tai tượng có khoảng cách di truyền rất hẹp.
Các kết quả phân tích chỉ thị phân tử rất cần thiết cho các nghiên cứu chọn giống và bố trí thí nghiệm của các vườn giống để tăng tỷ lệ thụ phấn chéo cho các cây trội có khoảng cách di truyền xa nhau nhằm tạo ra ưu thế lai ở đời sau.
Từ kết quả phân tích quan hệ di truyền của các dòng Keo tai tượng trên,
đề tài đã lựa chọn các cặp bố mẹ trong bố trí thí nghiệm vườn giống vô tính tại Yên Sơn - Tuyên Quang.
K82
K93
K116
K88
K102
K72
K110
K98
K91
K80
K109
K82
Hình 3.16: Minh họa sơ đồ bố trí các dòng Keo tai tượng trong vườn giống
Từ hình 3.15 đề tài đã minh họa sơ đồ bố trí các dòng Keo tai tượng trong vườn giống vô tính tại Yên Sơn - Tuyên Quang trong đó các dòng có quan hệ di truyền rất gần nhau như K93 - K98, K80 - K116, K72 - K88, K102 - K109, K91 - K110 và dòng K82 được bố trí xa nhau để hạn chế khả năng giao phấn giữa các dòng này với nhau, nhằm giảm thiểu thụ phấn cận huyết của những dòng có quan hệ di truyền quá gần nhau.