nghiên cứu cho thấy sự phát sinh rễ bất định thuận lợi hơn ở điều kiện sáng (phụ thuộc kiểu gen thực vật); ngoài ra, nhiều công bố cho thấy khi nuôi ở điều kiện sáng mạnh, ở rễ nhiều loài thực vật hình thành cấu trúc tương tự như lục lạp (chloroplast- like structure) có thể liên quan đến sự hình thành và tăng trưởng của rễ.
Đến nay đã ghi nhận nhiều nghiên cứu tạo rễ bất định với kết quả tốt với các điều kiện chiếu sáng khác nhau như tối - ví dụ trường hợp nuôi cấy mảnh lá Cichorium intybus [184], Centella asiatica [190], 40 µmol m-2 s-1 (3.000 lux) đối với Artemisia amygdalina [191], và 4.000 - 5.000 lux ở nuôi cấy tạo rễ từ mô lá Vernonia amygdalina [185]. Tương tự một số trường hợp nuôi cấy tạo rễ nêu trên, kết quả nghiên cứu này cho thấy mảnh lá NGBCC tái sinh rễ bất định tốt nhất ở nghiệm thức có cường độ chiếu sáng 4.000 lux.
3.1.1.4. Minh họa sự tái sinh rễ trực tiếp và khảo sát hình thái giải phẫu rễ tái sinh trực tiếp
Như đã trình bày, trong nghiên cứu này rễ bất định hình thành từ mô lá được định nghĩa theo cách tái sinh trực tiếp ngay ở lần nuôi cấy đầu tiên, khác với tái sinh gián tiếp qua trung gian giai đoạn mô sẹo.

Hình 3.44. Minh họa cận cảnh sự tái sinh trực tiếp rễ bất định từ mảnh lá nuôi cấy trên môi trường ½MS, 12 – 20 NSC.
A,B,C. Cận cảnh sự hình thành giai đoạn đầu của rễ đơn, cụm 2 rễ và cụm nhiều rễ ở 12 NSC; D,E,F. Sự phát triển tiếp theo của rễ đơn, cụm 2 rễ và cụm nhiều rễ, ở 20 ngày sau cấy.
Khá nhiều kết quả nghiên cứu về tái sinh trực tiếp rễ bất định từ mô lá đã được ghi nhận như tạo rễ Ophiorrhiza prostrata dùng môi trường ½MS với 10,74 µM NAA
110
và 2,32 µM kinetin [, tạo rễ Centella asiatica trên môi trường MS có 1 mg/L IBA [190], tạo rễ Rauwolfia serpentina trên môi trường MS có 1 mg/L PABA (para-amino benzoic acid) và 4 mg/L NAA [192]. Rễ Gynura procumbens tái sinh trực tiếp trên môi trường MS có bổ sung 3 mg/L IBA; ngược lại, ở môi trường MS có 3 mg/L NAA rễ tái sinh gián tiếp thông qua mô sẹo [182]. Ở NGBCC đây là kết quả nghiên cứu đầu tiên về tái sinh rễ trực tiếp từ vật liệu mảnh lá, được minh họa bằng các hình rễ tái sinh sau 12 đến 20 ngày nuôi cấy qua chụp cận cảnh (Hình 3.44).
Các sơ khởi rễ (giai đoạn ngày thứ 10 sau cấy) cũng đã được khảo sát hình thái giải phẫu với kết quả được trình bày dưới đây (Hình 3.45).
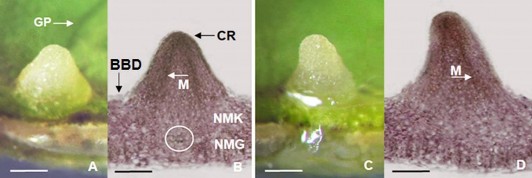
Hình 3.45. Hình thái giải phẫu sơ khởi rễ bất định hình thành trực tiếp từ mảnh lá. A,B. Sơ khởi rễ hình thành ở vị trí gân phụ của lá và hình thái giải phẫu tương ứng; C,D. Sơ khởi rễ hình thành không ở vị trí gân phụ của lá và hình thái giải phẫu tương ứng (hình chụp dưới kính hiển vi soi nổi với độ phóng đại 20X) (GP: Gân phụ của lá, BBD: Biểu bì mặt dưới của phiến lá, CR: Chóp rễ, M: vị trí bó mạch (cắt dọc), NMK: Nhu mô khuyết phiến lá, NMG: Nhu mô giậu phiến lá; vòng tròn chỉ vị trí mạch dẫn phụ của lá; thanh ngang 1 mm).
Sau 10 ngày nuôi cấy trên môi trường ½MS, dưới tác động của NAA (3 mg/L NAA) nhận thấy mép cắt và mô ở vị trí sát mép cắt của mẫu phiến lá phù ra, sơ khởi rễ hình thành trên bề mặt mẫu phiến lá gần mép cắt. Ở nghiên cứu này, có hai trường hợp khác nhau về vị trí tạo sơ khởi rễ đã được ghi nhận, một là, sơ khởi rễ hình thành ở ngay vị trí gân phụ của mẫu cấy với các cụm tế bào đang phân chia mạnh xuất phát từ các tế bào ở vị trí gần kề mạch (Hình 3.45A,B), hai là, sơ khởi rễ hình thành ở vị trí không có gân phụ - cũng ghi nhận có các cụm tế bào đang phân chia mạnh nhưng xuất phát từ các tế bào ở vị trí lục mô khuyết (Hình 3.45C,D). Ở trường hợp 1 sơ khởi rễ hình thành từ các tế bào liên kết với mô mạch, dưới tác động của NAA, các tế bào
111
này phân chia hình thành lớp tế bào tương tự tế bào tiền tượng tầng (procambial-like cells) đóng vai trò như tế bào gốc có nhiều tiềm năng (pluripotent stem cells) trong phân chia, phân hóa tạo sơ khởi rễ - tương tự kết quả nuôi cấy tạo rễ từ mảnh lá Medicago truncatula dưới tác động của auxin NAA [104], nuôi cấy tạo rễ từ mảnh lá Orthosiphon stamineus dưới ảnh hưởng của IAA /IBA/ NAA [193] và tạo rễ từ mảnh lá Labisia pumila dưới tác động của IBA/NAA [179]. Trước đó, Samaj và cộng sự (1999) cũng đã kết luận các lớp tế bào thuộc bó mạch (bundle sheet cells) chịu trách nhiệm phân chia, phân hóa tạo rễ trực tiếp ở Helianthus occidentalis [194]. Ở trường hợp 2, theo chúng tôi, sơ khởi rễ hình thành từ mô tế bào khuyết của mô thịt lá (spongy parenchyma/mesophyll cells) tương tự trường hợp nuôi cấy tái sinh chồi/rễ từ mảnh lá cây African violet (Sainpaulia ionantha) dưới ảnh hưởng của BA/ IBA [103]. Tái sinh rễ bất định ở hai trường hợp trên, theo chúng tôi, được giải thích là do các tế bào ở vị trí gần mạch và tế bào nhu mô khuyết, dưới tác động của sự di chuyển phân cực auxin đã được kích hoạt thay đổi lập trình di truyền dẫn đến tái sinh rễ [15].
3.3.2. Tạo rễ bất định từ chồi
3.3.2.1. Tạo rễ bất định từ chồi có nguồn gốc từ đốt thân cây vườn ươm
Bảng 3.21. Ảnh hưởng của NAA, IBA đến sự tạo rễ bất định từ chồi có nguồn gốc từ đốt thân cây vườn ươm, ở môi trường ½MS, 60 NSC.
Tỷ lệ chồi tạo rễ (%) | Số rễ/chồi | Chiều dài rễ (mm) | Đặc điểm hình thái, màu sắc rễ | |
ĐC | 100 | 4,33c* | 69,37c | Rễ mảnh, có rễ hơi nâu, ít lông hút, rễ nhánh dài |
NAA | 100 | 13,53a | 80,00a | Rễ có đường kính to, trắng, nhiều lông hút, rễ nhánh ngắn |
IBA | 100 | 11,67b | 74,56b | Rễ có đường kính trung bình, ít lông hút, rễ nhánh ngắn |
Có thể bạn quan tâm!
-

 Nhân Phôi Qua Nuôi Cấy Lỏng Lắc Trong Môi Trường Sh, 30 – 60 Nsc.
Nhân Phôi Qua Nuôi Cấy Lỏng Lắc Trong Môi Trường Sh, 30 – 60 Nsc. -
 Tạo Cây Con Từ Phôi Vô Tính Nuôi Lỏng Lắc Trong Môi Trường Ms, ½Ms
Tạo Cây Con Từ Phôi Vô Tính Nuôi Lỏng Lắc Trong Môi Trường Ms, ½Ms -
 Ảnh Hưởng Của Naa Và Môi Trường Khoáng Đến Sự Tạo Rễ Bất Định Trực Tiếp Từ Mảnh Lá (3 X 10 Mm), Ở Môi Trường ½Ms, 30 Nsc.
Ảnh Hưởng Của Naa Và Môi Trường Khoáng Đến Sự Tạo Rễ Bất Định Trực Tiếp Từ Mảnh Lá (3 X 10 Mm), Ở Môi Trường ½Ms, 30 Nsc. -
 Ảnh Hưởng Của Naa, Iba Đến Sự Phân Nhánh Rễ, Ở Môi Trường ½Ms, 21 Nsc.
Ảnh Hưởng Của Naa, Iba Đến Sự Phân Nhánh Rễ, Ở Môi Trường ½Ms, 21 Nsc. -
 Diễn Biến Tăng Trưởng Sinh Khối Rễ Bất Định Theo Thời Gian, 7 – 49 Nsc.
Diễn Biến Tăng Trưởng Sinh Khối Rễ Bất Định Theo Thời Gian, 7 – 49 Nsc. -
 C. Wu, Y.h. Duan, W. Tang, M.m. Li, X. Wu, G.c. Wang, W.c. Ye, G.x. Zhou, New Ursane-Type Triterpenoid Saponins From The Stem Bark Of Schefflera Heptaphylla, Fitoterapia, 2014, 92, 127-132.
C. Wu, Y.h. Duan, W. Tang, M.m. Li, X. Wu, G.c. Wang, W.c. Ye, G.x. Zhou, New Ursane-Type Triterpenoid Saponins From The Stem Bark Of Schefflera Heptaphylla, Fitoterapia, 2014, 92, 127-132.
Xem toàn bộ 180 trang tài liệu này.
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt ý nghĩa với p ≤ 0,05 trong phép thử LSD.
Các đoạn thân sau khử trùng được nuôi cấy trên môi trường MS có 0,5 mg/L BA và 2 mg/L kinetin. Kết quả cho thấy, dưới tác động kích thích của BA và kinetin, sau nuôi cấy từ 20 đến 45 ngày, chồi nách hình thành và phát triển, đến ngày thứ 60 sau cấy đạt kích thước (7 – 10 mm), xuất hiện 4 – 5 lá, có hình thái đặc trưng. Chồi
112
nách được tách ra cấy chuyển sang môi trường tạo rễ bất định, nhận thấy phần gốc mẫu cấy tiếp xúc với môi trường phù to do sùi mô sẹo, theo chúng tôi, đây là hiện tượng thường gặp trong nuôi cấy mô cây thân gỗ.
Các chồi in vitro được nuôi cấy tạo rễ bất định dùng môi trường ½MS có hoặc không bổ sung 0,2 mg/L IBA/NAA. Kết quả cho thấy trên môi trường không có NAA/IBA, rễ hình thành ít, rễ dài, có rễ màu hơi nâu, có ít lông hút, rễ nhánh dài (Hình 3.46A,D). Ở môi trường có IBA, chồi cũng tạo nhiều rễ, có ít lông hút, rễ nhánh ngắn (Hình 3.46B,E). Trên môi trường có NAA, chồi tạo rễ nhiều, rễ có đường kính to, dài, màu trắng, có nhiều lông hút, rễ nhánh cũng ngắn (Hình 3.46C,F). Kết quả (Bảng 3.19) cho thấy 100% chồi nuôi cấy ở tất cả các nghiệm thức đều tạo rễ bất định. Số rễ của chồi ở môi trường có NAA, IBA lần lượt là 13,53 và 11,67 so với ĐC là 4,3; chiều dài rễ tương ứng là 80 mm và 74,56 mm, theo thứ tự (so với ĐC 69,37 mm).

Hình 3.46. Tạo rễ bất định từ chồi có nguồn gốc từ đốt thân, 60 NSC. A,B,C. Cây 60 NSC ở môi trường ½MS (không có chất ĐHST); có 0,2 mg/L IBA; có 0,2 mg/L NAA. D,E,F. Rễ hình thành dưới đáy bình tương ứng của A,B,C. G, H. Cây hình thành ở môi trường ½MS có 0,2 mg/L IBA; 0,2 mg/L NAA. I. Cây từ phôi vô tính (thanh ngang 20 mm).
Đến nay, đã ghi nhận được một số công bố kết quả nghiên cứu dùng chồi (nguồn gốc từ thực địa) để tạo rễ bất định cây dược liệu, ví dụ, ngoài vật liệu mảnh lá, đoạn thân, Martin và cộng sự (2008) còn sử dụng chồi Ophiorrhiza prostrata (30
113
– 50 mm) nuôi cấy trên môi trường đặc MS có 10,74 µM NAA và 2,32 µM kinetin để tạo rễ, sau đó nuôi nhân thu sinh khối rễ nhằm nghiên cứu hợp chất thứ cấp camptothecin [131]. Yusuf và cộng sự (2018) cũng đã sử dụng chồi nụ (bud) Boesenbergia rotunda qua nuôi cấy trên môi trường đặc MS có 2 mg/L NAA để tạo rễ - như là nguồn vật liệu dùng nghiên cứu pinostrobin [195]. Rễ bất định loài sâm Java (Javanese ginseng) Talinum paniculatum được cảm ứng hình thành, thu thập từ chồi nuôi cấy trên môi trường đặc MS có 10 µM IBA, 30 g/L đường nhằm mục đích nghiên cứu sự tích lũy saponin [196].
3.3.2.2. Tạo rễ bất định từ chồi đốt thân cây in vitro
Sau cắt bỏ các lá, thân cây được cắt thành các đoạn thân mang chồi nách, do thân cây in vitro có đường kính nhỏ, các chồi nách lại khá gần nhau nên khó cắt chính xác các đoạn thân chỉ mang 1 chồi nách, do vậy ở ngày thứ 30 và 45 sau cấy nhận thấy một số mẫu cấy có 2 chồi phát triển. Sau nuôi cấy 60 ngày, các chồi được tách ra cấy sang môi trường tạo rễ bất định, nhận thấy thân chồi có đường kính nhỏ hơn thân chồi từ cây vườn ươm, phần thân trong môi trường nuôi cấy phù ra, mô sẹo hình thành nhưng ít hơn so với mẫu cấy cây vườn ươm, có thể do cây in vitro ở trạng thái “trẻ” hơn vì đã được nuôi cấy trong thời gian khá dài ở điều kiện in vitro.
Bảng 3.22. Ảnh hưởng của NAA và IBA đến sự tạo rễ bất định từ chồi đốt thân cây
in vitro, ở môi trường ½MS, 60 NSC.
Tỷ lệ chồi tạo rễ (%) | Số rễ/chồi | Chiều dài rễ (mm) | Đặc điểm hình thái, màu sắc rễ | |
ĐC | 100 | 3,84c* | 52,20c | Rễ mảnh, dài, trắng, ít lông hút, rễ nhánh dài |
NAA | 100 | 12,65a | 64,45a | Rễ có đường kính to, dài, trắng, nhiều lông hút, rễ nhánh ngắn |
IBA | 100 | 10,47b | 62,20b | Rễ có đường kính trung bình, dài, trắng, ít lông hút, rễ nhánh ngắn |
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt ý nghĩa với p
≤ 0,05 trong phép thử LSD.
Tương tự trường hợp tạo rễ từ chồi có nguồn gốc cây vườn ươm, kết quả tạo rễ cho thấy trên môi trường không có NAA/IBA (ĐC), rễ hình thành ít, rễ dài, ít lông
hút, rễ nhánh dài. Trên môi trường có IBA, chồi tạo nhiều rễ, rễ dài, ít lông hút, rễ nhánh ngắn (Hình 3.47B); trên môi trường có NAA, chồi tạo nhiều rễ, rễ có đường kính to, rễ dài, nhiều lông hút, rễ cũng ít phân nhánh (Hình 3.47C). Bảng 3.20 cho thấy 100% chồi nuôi cấy ở tất cả các nghiệm thức đều tạo rễ bất định. Số rễ của chồi ở môi trường có NAA, IBA lần lượt là 12,65 và 10,47 so với ĐC là 3,84; chiều dài rễ tương ứng là 64,45 mm và 62,20 mm, theo thứ tự (so với ĐC 52,20 mm). Nhận thấy số rễ, chiều dài rễ của chồi ở trường hợp này ít hơn, ngắn hơn so với rễ của chồi từ cây vườn ươm có thể do đường kính chồi từ cây in vitro nhỏ hơn – ảnh hưởng đến khả năng tạo rễ bất định.

Hình 3.47. Tạo rễ bất định từ chồi đốt thân cây in vitro, ở môi trường ½MS, 60 NSC
A. Thân cây in vitro cắt bỏ lá (vạch vàng vị trí cắt) B. Vật liệu tạo chồi nách C,D,E. Chồi nách phát triển ở 30 NSC, 40 NSC, 60 NSC (thanh ngang trắng 3 – 4 mm) F,G. Cây hình thành trên môi trường ½MS có 0,2 mg/L IBA; 0,2 mg/L NAA ở 60 NSC (thanh ngang 20 mm).
Cây/chồi in vitro là vật liệu tiền đề rất thuận lợi trong thực hiện thí nghiệm vì đây là nguồn mẫu sẵn ở trạng thái vô trùng, có thể thu các loại vật liệu khác nhau như chồi ngọn/chồi nách, lá, thân, rễ dùng trực tiếp trong nhiều nghiên cứu khác nhau về nhân giống, sự tích lũy hợp chất thứ cấp. Các chồi, có nguồn gốc chồi nách cây in vitro Ophiorrhiza mungos qua nuôi cấy trên môi trường MS có 0,25 mg/L BA và 025 mg/L kinetin, tạo rễ bất định (trên môi trường ½MS có 100 mg/L than hoạt tính) thành cây hoàn chỉnh được sử dụng trong nghiên cứu hợp chất thứ cấp kháng ung thư camptothecin [197].
3.3.2.3. Tạo rễ bất định từ chồi có nguồn gốc phôi vô tính
Cây con từ phôi vô tính (Hình 3.48A), sau khi cắt bỏ phần rễ trụ, phần thân mang các lá được cấy vào môi trường ½MS có chứa 0,1 mg/L NAA/IBA để tạo rễ bất định. Sau 40 ngày nuôi cấy, trên môi trường không có chất điều hòa sinh trưởng (NAA/IBA), chồi tạo rễ khá tốt, rễ ít lông hút, có số rễ/chồi là 3,13 với chiều dài rễ trung bình là 25,67 mm (Hình 3.48B); môi trường nuôi cấy có 0,1 mg/L IBA, rễ dài, ít lông hút, có rễ nhánh; môi trường có 0,1 mg/L NAA rễ hình thành nhiều, cũng có hình thành rễ nhánh, nhưng rễ có nhiều lông hút đây là sự khác biệt khác biệt so với rễ hình thành ở môi trường có IBA, (Hình 3.48C,D). Như vậy, ở môi trường tạo rễ có bổ sung NAA/IBA hoặc không có chất ĐHST có ảnh hưởng khác biệt lên sự tạo rễ, NAA hiệu quả hơn IBA.
Bảng 3.23. Ảnh hưởng của NAA và IBA đến sự tạo rễ bất định từ chồi có nguồn gốc phôi vô tính, môi trường ½MS, 30 NSC.
Tỷ lệ chồi tạo rễ (%) | Số rễ/chồi | Chiều dài rễ (mm) | Đặc điểm hình thái, màu sắc rễ | |
ĐC | 100 | 3,13c* | 25,67b | Rễ dài, trắng, ít lông hút, không có rễ nhánh |
IBA | 100 | 4,37a | 37,67a | Rễ dài, trắng, ít lông hút, có rễ nhánh |
NAA | 100 | 6,33b | 39,78b | Rễ dài, trắng, nhiều lông hút, gốc rễ to, có rễ nhánh |
*Các chữ cái khác nhau trong một cột thể hiện sự khác biệt ý nghĩa với p ≤ 0,05 trong phép thử LSD.
Phôi nói chung, phôi vô tính hoặc phôi hạt nói riêng và dạng phát triển tiếp theo của chúng (cây con) cũng là nguồn vật liệu được các nhà nghiên cứu sử dụng trong một số trường hợp, ví dụ như trường hợp tạo phôi vô tính từ nuôi cấy mảnh lá mầm phôi hạt sâm Mỹ Panax quinquefolius trên môi trường MS có 50 g/L đường, tạo rễ trụ thành cây hoàn chỉnh trên môi trường 1/3SH, có bổ sung 0,25 mg/L NAA, 1% sucrose với mục đích nhân giống [36]. Baskaran và Jayabalan (2009) đã dùng đoạn mô trụ dưới lá mầm (10 mm) cây từ phôi hạt Psoralea coryfolia (10 ngày tuổi) nuôi cấy trên môi trường MS có bổ sung 4 µM IAA, NAA để tạo rễ bất định và sau
đó rễ bất định được nuôi nhân thu sinh khối trong môi trường ½MS có 3 µM IBA dùng nghiên cứu hàm lượng psoralen [198].

Hình 3.48. Tạo rễ bất định từ chồi có nguồn gốc phôi vô tính, ở môi trường ½MS, 30 NSC.
A. Vật liệu tạo rễ bất định. Rễ bất định hình thành ở môi trường không có chất điều hòa sinh trưởng (B); có 0,1 mg/L IBA (C); 0,1 mg/L NAA (D), (thanh ngang 10mm).
Như vậy, ở NGBCC, chồi cũng là vật liệu khá thuận lợi trong tạo rễ bất định và NAA là tác nhân rất thích hợp trong kích thích tạo rễ từ chồi. Kết quả tác động của NAA trong nghiên cứu này phù hợp với kết quả đáp ứng tích cực đối với tạo rễ của mô lá trong điều kiện nuôi cấy có NAA như đã trình bày ở trên. Sự tác động của chất điều hòa sinh trưởng thực vật lên mô lá, mô thân có sự khác biệt rất rõ nét, với mô lá hoàn toàn không có biểu hiện cảm ứng với IBA ở các nồng độ khác nhau 0,5; 1; 2; 3; 4; 5 mg/L trên các môi trường nuôi cấy cơ bản là MS; ½MS; B5; SH. Nhưng với các thí nghiệm sử dụng vật liệu chồi, IBA có tác động gây cảm ứng tạo rễ bất định trên môi trường có ½MS. Điều đó chứng tỏ rằng, sự tác động của mỗi loại chất điều hòa sinh trưởng là khác nhau và trên từng loại mô là khác nhau. Đồng thời, vật liệu mô lá cho kết quả tạo rễ hiệu quả hơn so với vật liệu từ chồi, do vậy, chỉ rễ bất định từ mô lá mới được sử dụng trong nhân rễ.
Akhami và cộng sự (2013) [101] đã ghi nhận hiện tượng IAA tăng lên ở vị trí gốc đoạn thân mang chồi Petunia hybrida (trồng trong khay nhựa) ở thời điểm 2 h và 24 h sau cắt đoạn và kết luận sự phát sinh rễ bất định phụ thuộc vào đỉnh IAA ở thời điểm 24 h và sự di chuyển phân cực của auxin - cảm ứng sự phân chia đầu tiên của tế bào meristem dẫn đến tạo sơ khởi rễ, IAA giảm xuống sau thời điểm 24 h tạo thuận lợi cho quá trình phát triển tiếp theo của rễ. Guan và cộng sự (2019) [101] cho rằng rễ bất định ở đoạn thân mang chồi 2 dòng cà chua (dòng Alisa Craig và dòng chuyển gen DR5pro:YFP trans, nuôi trong dung dịch dinh dưỡng Hoagland) bắt nguồn từ tế