20.00
20.00
18.00
17.42b
16.12d
18.26a
16.91c
18.47a
17.07
Có thể bạn quan tâm!
-

 Ảnh Hưởng Của Naa Và Môi Trường Khoáng Đến Sự Tạo Rễ Bất Định Trực Tiếp Từ Mảnh Lá (3 X 10 Mm), Ở Môi Trường ½Ms, 30 Nsc.
Ảnh Hưởng Của Naa Và Môi Trường Khoáng Đến Sự Tạo Rễ Bất Định Trực Tiếp Từ Mảnh Lá (3 X 10 Mm), Ở Môi Trường ½Ms, 30 Nsc. -
 Minh Họa Sự Tái Sinh Rễ Trực Tiếp Và Khảo Sát Hình Thái Giải Phẫu Rễ Tái Sinh Trực Tiếp
Minh Họa Sự Tái Sinh Rễ Trực Tiếp Và Khảo Sát Hình Thái Giải Phẫu Rễ Tái Sinh Trực Tiếp -
 Ảnh Hưởng Của Naa, Iba Đến Sự Phân Nhánh Rễ, Ở Môi Trường ½Ms, 21 Nsc.
Ảnh Hưởng Của Naa, Iba Đến Sự Phân Nhánh Rễ, Ở Môi Trường ½Ms, 21 Nsc. -
 C. Wu, Y.h. Duan, W. Tang, M.m. Li, X. Wu, G.c. Wang, W.c. Ye, G.x. Zhou, New Ursane-Type Triterpenoid Saponins From The Stem Bark Of Schefflera Heptaphylla, Fitoterapia, 2014, 92, 127-132.
C. Wu, Y.h. Duan, W. Tang, M.m. Li, X. Wu, G.c. Wang, W.c. Ye, G.x. Zhou, New Ursane-Type Triterpenoid Saponins From The Stem Bark Of Schefflera Heptaphylla, Fitoterapia, 2014, 92, 127-132. -
 Nghiên cứu tạo, nhân phôi vô tính và rễ bất định cây ngũ gia bì chân chim Schefflera octophylla Lour. Harms - 21
Nghiên cứu tạo, nhân phôi vô tính và rễ bất định cây ngũ gia bì chân chim Schefflera octophylla Lour. Harms - 21 -
 Nghiên cứu tạo, nhân phôi vô tính và rễ bất định cây ngũ gia bì chân chim Schefflera octophylla Lour. Harms - 22
Nghiên cứu tạo, nhân phôi vô tính và rễ bất định cây ngũ gia bì chân chim Schefflera octophylla Lour. Harms - 22
Xem toàn bộ 180 trang tài liệu này.
bc
16.00
16.00
13.92e
14.00
12.83f
.04a
.19
a
2.44b
12.00
12.00
12.08c 12.19bc
10.00
9.94e
11.50
9.16
f
8.00
7.90g
8.00
6.00
5.13i
4.00
4.52j
3.66i
3.23j
7.15h
5.64g
5.11h
4.00
2.00
2.62k
1.8
2.18l
56l
0.00
0.00
7 14 21
28
Thời gian (ngày)
35
42
49
Khối lượng rễ thu nhận (NAA)
Hệ số nhân sinh khối (NAA)
Khối lượng rễ thu nhận (IBA)
Hệ số nhân sinh khối (IBA)
7k
1.
13
d
13
1
Khối lượng rễ thu nhận (g)
Hệ số nhân sinh khối (lần)
so với khối lượng tươi rễ thu nhận ở 42 NSC (do đã ở thời kỳ ổn định); ngoài ra, nhận thấy ở giai đoạn 49 NSC, một số rễ có hiện tượng hóa nâu (Hình 3.55) có thể do hàm lượng dinh dưỡng môi trường đã giảm nhiều. Tương tự, rễ nuôi trong môi trường IBA cũng có xu hướng tăng sinh khối rất nhanh ở 28 NSC và đạt mức tối đa ở 42 NSC (Hình 3.54G,H) (khối lượng tươi rễ lần lượt là 12,83 g và 16,91 g; hệ số nhân tương ứng là 9,16 và 12,08), đến giai đoạn 49 NSC nhận thấy khối lượng rễ tươi thu nhận được không có sự khác biệt về mặt thống kê so với thời điểm 42 NSC. Rễ nuôi trong môi trường có NAA phát triển nhanh hơn về khối lượng ở từng thời điểm và từng nồng độ so với rễ trong môi trường có IBA (Hình 3.54G,H).
Biểu đồ 3.1. Diễn biến tăng trưởng sinh khối rễ bất định theo thời gian, 7 – 49 NSC.
Nhằm thu sinh khối ở mức tối ưu dùng cho các mục đích khác nhau, đối với bất kỳ hệ thống nuôi cấy (lỏng lắc, bioreactor) và vật liệu nuôi cấy nào (tế bào, phôi, rễ,..), khảo sát động học tăng trưởng theo thời gian của vật liệu nuôi cấy là yêu cầu quan trọng. Trong điều kiện nuôi lỏng lắc, nhiều tài liệu cho thấy giai đoạn rễ đạt giá trị khối lượng tối đa ở một thời điểm nuôi cấy nhất định tùy thuộc vào loài thực vật, cụ thể Ahmad và cộng sự (2020) ghi nhận 3 pha tăng trưởng của rễ bao gồm pha mở đầu (7 – 9 ngày), pha tăng trưởng rất nhanh (12 - 27 ngày) và đạt mức ổn định từ 27
– 30 ngày qua nghiên cứu nuôi rễ bất định Stevia rebaudiana [204]; cũng qua nghiên cứu sự phát triển sinh khối theo thời gian (từ 7 – 49 ngày) của rễ bất định Prunellavulgaris, Fazal và cộng sự (2014) cho thấy khối lượng tươi rễ đạt mức cao nhất chỉsau 21 ngày nuôi [196]. Sinh khối rễ Psoralea corylifolia đạt mức tối đa sau 4 tuần nuôi cấy [198]; cần 5 tuần đối với rễ Panax notoginseng [44], Vernonia amygdalina [185], 6 tuần đối với rễ chicory (Cichorium intybus) tuy nhiên, rễ Cichorium intybus mất dần khả năng tạo rễ nhánh khi duy trì nuôi cấy dài hạn [184].
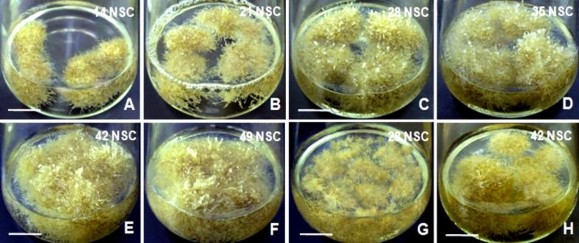
Hình 3.54. Tăng trưởng sinh khối rễ bất định theo thời gian.
A,B. Sinh khối rễ trong môi trường có 2 mg/L NAA ở 14 NSC, 21 NSC; C,D. Sinh khối rễ trong môi trường có 2 mg/L NAA ở 28 NSC, 35 NSC; E,F. Sinh khối rễ trong môi trường có 2 mg/L NAA ở 42 NSC, 49 NSC; G,H. Sinh khối rễ trong môi trường có 2 mg/L IBA ở 28 NSC, 42 NSC, theo thứ tự (thanh ngang 2 cm).
Nói chung, rễ NGBCC đã thể hiện các pha tăng trưởng trong nuôi cấy như pha mở đầu, tăng trưởng rất nhanh, pha ổn định và cần thời lượng 5 – 6 tuần để đạt mức khối lượng tươi rễ cao nhất (Biểu đồ 3.1) - phù hợp với nghiên cứu nuôi rễ của một số tác giả nêu trên. Qua nuôi cấy dài hạn rễ NGBCC, không ghi nhận được trường hợp hóa thuỷ tinh thể dẫn đến mô ở trạng thái bị ‘trong’, có thể hoại tử sau đó – hiện tượng sinh lý thường gặp ở nuôi cấy mẫu ngập một phần/toàn phần (thường là chồi) trong môi trường lỏng [205]; cũng nhận thấy rễ không giảm khả năng phát sinh rễ thứ cấp, tạo tiền đề thuận lợi cho thử nghiệm nuôi nhân rễ ở quy mô lớn hơn.
KẾT LUẬN VÀ KIẾN NGHỊ
KẾT LUẬN
Trên cơ sở ứng dụng phương pháp nuôi cấy mô tế bào thực vật, lần đầu tiên trên đối tượng NGBCC, đã xác định được:
Sự phát sinh phôi vô tính trực tiếp từ mô lá hiệu quả nhất ở môi trường đặc SH có bổ sung 5 mg/L NAA, 0,25 mg/L BA, 50 g/L sucrose, 10% nước dừa; điều kiện chiếu sáng 4.000 lux. Phát sinh phôi gián tiếp qua mô sẹo mảnh lá ở môi trường đặc SH có 2 mg/L NAA, 0,25 mg/L BA, 30 g/L sucrose.
Môi trường nuôi cấy lỏng SH có bổ sung 2 mg/L NAA, 0,25 mg/L BA, 50 g/L sucrose, 10% nước dừa ở điều kiện chiếu sáng 4.000 lux là tối ưu cho sự phát sinh phôi thứ cấp, với khối lượng phôi nuôi cấy 2% và kích thước phôi nuôi cấy ~ 10 mm là hiệu quả nhất. Phôi thứ cấp hình thành trong môi trường lỏng lắc hiệu quả hơn so với môi trường đặc.
Sự tăng trưởng của cây con từ phôi vô tính hiệu quả nhất ở môi trường ½MS (20 g/L sucrose), cây con sinh trưởng bình thường ở điều kiện ex vitro.
Rễ bất định phát sinh trực tiếp từ mô lá hiệu quả cao nhất trên môi trường đặc
½MS có bổ sung 3 mg/L NAA, 30 g/L sucrose, điều kiện chiếu sáng 4.000 lux.
Thu nhận sinh khối rễ bất định thông qua sự tạo rễ thứ cấp bằng phương pháp nuôi lỏng lắc trong môi trường ½MS có bổ sung 2 mg/L NAA, 30 g/L sucrose; khối lượng rễ nuôi cấy ban đầu thích hợp là 2% đạt hiệu quả cao nhất.
Rễ bất định phát sinh từ chồi đốt thân (cây vườn ươm/in vitro) nuôi cấy trên môi trường thích hợp ½MS có 0,2 mg/L NAA có 20 g/L sucrose. Chồi có nguồn gốc phôi vô tính tạo rễ hiệu quả qua nuôi cấy trên môi trường đặc ½MS có 0,1 mg/L NAA (20 g/L sucrose).
Khảo sát hình thái giải phẫu phôi vô tính (ở các giai đoạn phát triển khác nhau) và rễ bất định bằng phương pháp nhuộm kép đã được thực hiện có kết quả.
Các kết quả nghiên cứu trên góp phần phát triển hướng nghiên cứu về cơ sở sinh lý của tính toàn thế của tế bào thực vật nói chung, ở đối tượng Schefflera octophylla nói riêng.
KIẾN NGHỊ
Nghiên cứu nuôi nhân phôi vô tính, rễ bất định ở quy mô lớn bằng hệ thống bioreactor.
Sử dụng sản phẩm sinh khối phôi vô tính (từ kết quả nhân phôi theo cơ chế tạo phôi thứ cấp), rễ bất định trong nghiên cứu nhân giống, thu nhận các hợp chất thứ cấp phục vụ lĩnh vực y dược/mỹ phẩm và trong một số nghiên cứu quan trọng khác như bảo quản nguồn gen, biến nạp gen,...
DANH MỤC CÔNG TRÌNH
1. Huỳnh Thị Lũy, Nguyễn Hữu Hổ, Bùi Văn Lệ, Tạo phôi vô tính trực tiếp từ nuôi cấy in vitro mô lá cây Ngũ gia bì chân chim Schefflera octophylla (Lour.) Harms, Kỷ yếu Hội nghị Công nghệ Sinh học toàn quốc, TP. HCM, 2019, 405-410.
2. Huỳnh Thị Lũy, Nguyễn Hữu Hổ, Bùi Văn Lệ, Tạo rễ bất định trực tiếp từ mô lá cây Ngũ gia bì chân chim Schefflera octophylla (Lour.) Harms nuôi cấy in vitro, Tạp chí Công nghệ Sinh học, 2021, 19(3), 495-507.
3. Huỳnh Thị Lũy, Nguyễn Hữu Hổ, Bùi Văn Lệ, Nhân rễ bất định cây Ngũ gia bì chân chim Schefflera octophylla (Lour.) Harms bằng phương pháp nuôi lỏng lắc, Tạp chí Nông nghiệp và Phát triển Nông thôn, 2021, 6(3), 12-20.
TÀI LIỆU THAM KHẢO
1. Lã Đình Mỡi, Châu Văn Minh, Trần Văn Sung, Phạm Quốc Long, Phan Văn Kiệm, Trần Huy Thái, Trần Minh Hợi, Ninh Khắc Bản, Lê Mai Hương, Họ Nhân sâm (Araliaceae Juss.) - Nguồn hoạt chất sinh học đa dạng và đầy triển vọng ở Việt Nam, Tuyển tập Hội nghị khoa học toàn quốc lần thứ 5 về Sinh thái và Tài nguyên Sinh vật, Viện Hàn Lâm Khoa học và Công nghệ Việt Nam, 2013, 1152-1158.
2. J. Kitajima, M. Shindo, Y. Tanaka, Two new triterpenoid sulfates from the leaves of Schefflera octophylla, Chemical Pharmaceutical Bulletin, 1990, 38(3), 714-716.
3. Y.L. Li, D.X. Shi, M.L. Wang, Tissue culture and rapid propagation of Schefflera octophylla, Plant Physiology Communications, 2004, 40(2), 193.
4. Y.L. Li, P.P. But, V.E.C. Ooi, Antiviral activity and mode of action of caffeoylquinic acids from Schefflera heptaphylla (L.) Frodin, Antiviral Research, 2005, 68(1), 1-9.
5. Y.L. Li, R.W. Jiang, L.M.S. Ooi, P.P.H. But , V.E.C. Ooi, Antiviral triterpenoids from the medicinal plant Schefflera heptaphylla. Phytother Research, 2007, 21(5), 466-470.
6. Y.L. Li, C.M. Yeung, L.C.M. Chiu, Y.Z. Cen, V.E.C. Ooi, Chemical composition and antiproliferative activity of essential oil from the leaves of a medicinal herb, Schefflera heptaphylla, Phytother Research, 2009, 23(1), 140- 142.
7. Y.F. Chen, S.H. Tao, F.L. Zeng, L.W. Xie, Z.B. Shen, Antinociceptive and anti- inflammatory activities of Schefflera octophylla extracts, Journal Ethnopharmacology, 2015, 171, 42-50.
8. X. Liu, Y. Niu, J. Liu, R. Xu, M. Shi, W. Kang, Efficient extraction of anti- inflammatory active ingredients from Schefflera octophylla leaves using ionic liquid-based ultrasonic-assisted extraction coupled with HPLC, Molecules, 2019a, 24(16), 2942.
9. X. Liu, J. Dong, Q. Liang, H.M.D. Wang, X.H. Liu, Coagulant effects and mechanism of Schefflera heptaphylla (L.) Frodin, Molecules, 2019b, 24(24), 4547.
10. Đỗ Huy Bích, Đặng Quang Chung, Bùi Xuân Chương, Nguyễn Thượng Dong, Đỗ Trung Đàm, Phạm Văn Hiền, Vũ Ngọc Lộ, Phạm Duy Mai, Phạm Kim Mẫn, Đoàn Thị Thu, Nguyễn Tập, Trần Toàn, Cây thuốc và động vật làm thuốc ở Việt Nam, Nhà xuất bản Khoa học và Kỹ thuật, Viện Dược liệu, 2006, 2, 411-414.
11. Đặng Thị Thanh Tâm, Nghiên cứu tạo rễ tơ của cây Tam thất, Ngũ gia bì chân chim và thử nghiệm quá trình sản xuất sinh khối bằng bioreactor, Báo cáo tổng hợp đề tài KC.04.TN10/11-15 thuộc Chương trình KH&CN trọng điểm cấp nhà nước (KC.04/11-15), Trường Đại học Nông nghiệp Hà Nội, 2012.
12. W. Gaoyin, W. Xiaoli, W. Xiao, L. Xian, W. Yi, Changes in biochemistry and histochemical characteristics during somatic embryogenesis in Ormosia henryi Prain, BioRxiv, 2020; https://doi.org/10.1101/2020.09.21.307009.
13. L. Guan, Y. Li, K. Huang, Z.M. Cheng, Auxin regulalation and MdPIN expression during adventitious root initiation in apple cuttings, Hoticulture Reseach, 2020, 7: 143.
14. S.K.Verma, A.K.Das, S.Gantait, S. Gurel, E. Gurel, Influence of auxxin and its polar transport inhibitor on the development of somatic embryos in Digitalis trojana, 2018, 8 (2), 99.
15. Y.H. Su, L.P. Tang, X.Y. Zhao, X.S. Zhang, Plant cell totipotency: Insights into cellular reprogramming, Plant Biology Journal of Integrative, 2021, 63, 228– 240.
16. N. Vidal, C. Sanchez, Use of bioreactor systems in the propagation of forest trees, Engineering in Life Sciences, 2019, 19 (12), 896-915.
17. B. Qiang, J. Miao, N. Phillips, K. Wei, Y. Gao, Recent advances in the tissue culture of American ginseng (Panax quinquefolius), Chemistry and Biodiversity, 2020, 17(10), e2000366.
18. F. Engelmann, In vitro conservation of tropical plant germplasm - a review, Euphytica, 1991, 57, 227-243.
19. Y. Guan, S.G. Li, X.F. Fan, Z.H. Su, Application of somatic embryogenesis in woody plants, Front Plant Science, 2016, 7, 938.
20. V. Pikulthong, T. Teerakathiti, A. Thamchaipenet, S. Peyachoknagul,
Development of somatic embryos for genetic transformation in Curcuma longa
132
L. and Curcuma mangga Valeton & Zijp. Agriculture and Natural Resources, 2016, 50, 276-285.
21. F. Afreen, S.M.A. Zobayed, T. Kozai, Photoautotrophic culture of Coffea arabusta somatic embryos: Development of a bioreactor for large-scale plantlet conversion from cotyledonary embryos. Annals of Botany, 2002, 90, 21-29.
22. H. Etienne, D. Breton, J.C. Breitler, B. Bertrand, E. Déchamp, R. Awada, P. Marraccini, S. Léran, E. Alpizar, C. Campa, P. Courte, F. Georget, J.P. Ducos, Coffee somatic embryogenesis: How did research, experience gained and innovations promote the commercial propagation of elite clones from the two cultivated species? Frontiers in Plant Science, 2018, 9, 01630.
23. Nguyễn Trung Thành và Paek Kee Yoeup, Nhân nhanh rễ bất định Nhân sâm Panax ginseng C. A. Meyer: Ảnh hưởng của một số nhân tố lý hóa lên sự tăng trưởng sinh khối và sản phẩm trao đổi chất ginsenosides, Tạp chí Khoa học ĐHQGHN, Khoa học Tự nhiên và Công nghệ, 2008, 24, 318-323.
24. Dương Tấn Nhựt, Trần Hiếu, Nguyễn Thị Nhật Linh, Hoàng Thanh Tùng, Nguyễn Bá Nam, Nguyễn Phúc Huy, Vũ Quốc Luận, Vũ Thị Hiền. Tối ưu hóa quá trình nhân nhanh và tích lũy saponin của rễ bất định sâm Ngọc Linh (Panax vietnamensis Ha et Grushv.) trong các hệ thống nuôi cấy, Tạp chí Công nghệ Sinh học, 2015, 13(3), 853-864.
25. Y. Wang, S. Jia, Z. Wang, Y. Chen, S. Mo, N.N. Sze, Planning considerations of green corridors for the improvement of biodiversity resilience in suburban areas, Journal of Infrastructure Preservation and Resilience, 2021, 2:6.
26. H.G.A. Engler and K.A.E. Prantl, Die Natürlichen Pflanzenfamilien, 1894, 3(8): 38, Angelmann Publisher.
27. Tự điển Bách khoa Dược học, Nhà xuất bản Từ điển Bách khoa Hà Nội, 1999, 433-434.
28. Trần Văn Sung, G. Adam, A sulphated triterpenoid saponin from Schefflera octophylla. Phytochemistry, 1991, 30 (8), 2717-2720.
29. Trần Văn Sung, J. Peter, Katalinic, G. Adam, A bisdesmosidic triterpenoid saponin from Schefflera octophylla, Phytochemistry, 1991a, 30 (11), 3717- 3720.
30. Trần Văn Sung, Steglich, G. Adam, Triterpenoid glycosides from Schefflera