thực vật. Hằng năm, có một lượng lớn cellulose từ các mảng thực vật khô hình thành và chiếm từ 35% - 50% trữ lượng cellulose [24].
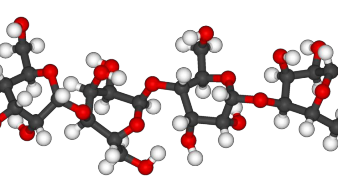
Hình 1.5 Cấu trúc không gian của phân tử cellulose [9]
Các gốc D- glucose trong cellulose thường lệch nhau một góc 1800 và có dạng như một chiếc ghế bành [45].
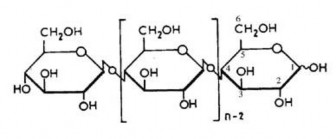
Hình 1.6.Cấu trúc hóa học phân tử cellulose [9]
Hemicellulose
Hemicellulose cũng là các đại phân tử nhưng mạch cấu tạo từ các đường 5 cacbon pentose và các đường 6 cacbon hexose tuy nhiên đường 5 cacbon là chủ yếu. Hemicellulose tạo liên kết với cellulose tạo nên cấu trúc thành tê bào thực vật.
Có thể bạn quan tâm!
-

 Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 2
Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 2 -
![Đặc Tính Của Cellulose Và Xylanase Từ C.cellulans [18]](https://tailieuthamkhao.com/uploads/2023/02/19/nghien-cuu-cellulase-tu-vi-khuan-ruot-moi-phan-lap-o-viet-nam-3-1-120x90.jpg) Đặc Tính Của Cellulose Và Xylanase Từ C.cellulans [18]
Đặc Tính Của Cellulose Và Xylanase Từ C.cellulans [18] -
![Tính Bền Nhiệt Của Cellulase Và Xylanase Từ C.cellulans [18].](https://tailieuthamkhao.com/uploads/2023/02/19/nghien-cuu-cellulase-tu-vi-khuan-ruot-moi-phan-lap-o-viet-nam-4-1-120x90.jpg) Tính Bền Nhiệt Của Cellulase Và Xylanase Từ C.cellulans [18].
Tính Bền Nhiệt Của Cellulase Và Xylanase Từ C.cellulans [18]. -
 Phương Pháp Phân Lập Và Tuyển Chọn Chủng Sinh Tổng Hợp Cellulase
Phương Pháp Phân Lập Và Tuyển Chọn Chủng Sinh Tổng Hợp Cellulase -
 Khảo Sát Khả Năng Thủy Phân Rơm Sử Dụng Cellulase Thu Nhận Từ Vi Khuẩn
Khảo Sát Khả Năng Thủy Phân Rơm Sử Dụng Cellulase Thu Nhận Từ Vi Khuẩn -
 Phương Pháp Xác Định Lượng Đường Trong Mẫu Thủy Phân Bằng Sắc Ký Lỏng Cao Áp (Hplc- High Performance Liquid Chromatography)
Phương Pháp Xác Định Lượng Đường Trong Mẫu Thủy Phân Bằng Sắc Ký Lỏng Cao Áp (Hplc- High Performance Liquid Chromatography)
Xem toàn bộ 138 trang tài liệu này.
Về thành phần, hemicellulose bao gồm các polysaccharid như xylan (22.5%), arabinan (1.3%), galactan (1.3%) và mannan (3.4%). Các polysaccharid này được hình thành từ các đường đơn như D-xylose, D-mannose, D-glucose, D-galactose, L- arabinose. Do hemicellulose trong bã mía chứa nhiều đường xylose, nên thường được sử dụng để sản xuất xylitol [79]. Đối với các loại gỗ mềm và các loại cỏ, bèo
…, phần lớn đường pentose trong hemicellulose là mannose, trong khi đó đối với gỗ cứng là đường 5 xylose. Do mạch hemicellulose ngắn hơn và có cả mạch nhánh nên so với cellulose thì hemicellulose dễ bị thủy phân hơn [46].
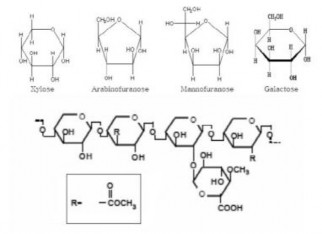
Hình 1.7 Công thức hóa học của hemicellulose [29]
Lignin
Thành phần thứ ba của lignocellulose là lignin. Lignin là một polyme dạng vòng, có cấu trúc bậc 3 của mạng polyphenol thơm, bao gồm các đơn phân phenylpropannoid, dẫn xuất của n-hydroxycinnamyl alcohol được dimethoxyl hóa (syringyl), monomethoxyl hóa (guaiacyl) hoặc không được methoxyl hóa (p- hydroxyphenil) [47]. Lignin là phân tử kỵ nước và bền với các tác nhân phân giải hóa học và sinh học, đóng vai trò như lớp kết dính giữa các tế bào thực vật và các lớp màng tế bào, tạo với hemicellulose và cellulose một kết cấu trong đó các sợi cellulose được bảo vệ khỏi các tác nhân bên ngoài. Lignin liên kết với các thành phần polysaccharid của lignocellulose bằng các liên kết cộng hóa trị bởi nhiều dạng liên kết etherbenzylic, esterbenzyle, glycoside và acetyle [48].
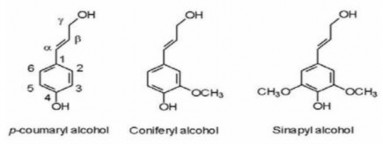
Hình 1.8 Các đơn phân của lignin
Sinh khối tạo thành từ Lignocellulose chiếm khoảng 50% tổng lượng sinh khối thực vật trên thế giới với ước tính trong khoảng 10-50 tỷ tấn [44].
1.3.2. Hệ enzym phân giải Lignocellulose
Ngoài các enzym thuộc nhóm cellulase tham gia vào quá trình thủy phân lignocellulose còn có hệ hemicellulase tham gia đồng thời vào quá trình phân giải các sinh khối thực vật. Hemicellulase gồm một nhóm các enzym thủy phân các galactan, xylan, mannans và araban. Các enzym chính trong hệ hemicellulase bao
gồm: Endoxylanase (EC3.2.1.8, 1,4-β-d xylan xylanohydrolase), enzym này thủy phân liên kết β-d xylano pyranosyl của xylan thành các dạng xylo-oligosaccharide; β-D xylosidase (EC 3.2.1.3, xylobiase) xúc tác thủy phân các xylobiose hoặc xylo- oligosaccharide từ đầu không khử, sản phẩm tạo thành là các đường D-xylose [49].
Cellulase và Hemicellulase là các glycoside hydrolase (GHs), chúng phân cắt các liên kết glycoside giữa hai hay nhiều hơn các carbohydrate hoặc giữa các carbohydrate với nửa không phải carbohydrate. GHs được phân loại dựa trên trình tự protein và các chuỗi peptid, nó không có hoạt tính đặc hiệu. Hiên nay có 133 họ GHs. Ngoài họ GHs, có nhiều enzym carbohydrate khác có hoạt tính phân cắt cellulose và hemicellulose. Các enzym này được chia làm 3 nhóm: polysaccharide lyases (PLs), carbohydrate esterases (CEs) và họ enzym hoạt hóa bổ trợ (AAs). Một nhóm các protein quan trọng khác có sự liên kết với các enzym hoạt hóa carbohydrate đó là các carbohydrate binding modules (CBMs). Có khoảng 68 họ CBM được liệt kê trong CAZy, cũng được phân loại bởi trình tự và cấu trúc gấp nếp, chúng nhận dạng các tinh thể cellulose, các cellulose vô định hình và cấu tạo của hemicellulose cũng như các carbonhydrate khác. CBMs đóng vai trò quan trọng trong việc khoanh vùng chính xác để enzym carbohydrate tấn công cơ chất [50] Endoglucanase của nấm mốc quá trình xúc tác nói chung không tạo ra các carbohydrate –binding modules (CBM) trong khi đó các endoglucanase của vi khuẩn có thể tạo ra các CBM [11].
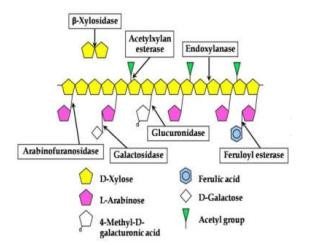
Hình 1.9 Hoạt động của các enzym hemicellulose [51]
1.3.3. Sự phối hợp tác động giữa các enzym
Từ những nghiên cứu riêng lẻ đối với từng loại enzym đến nghiên cứu tác động tổng hợp của cả ba loại enzym, nhiều nhà khoa học đều đưa ra kết luận chung là các enzym cellulase sẽ thay nhau phân hủy cellulose để tạo thành sản phẩm cuối
cùng là glucose. Để phân hủy được cellulose, trước hết bề mặt cũng như các phần cấu trúc của cellulose cần phải được nới lỏng giúp các enzym có thể xâm nhập và tiếp xúc trực tiếp với các sợi cellulose ở bên trong. Một khi các cellulase xâm nhập vào được mạng lưới cellulose sẽ từng bước tiến hành phân hủy cellulose để tạo thành sản phẩm cuối cùng là D-glucose. Sự thủy phân hoàn toàn cellulose thành sản phẩm cuối cùng glucose cần có hoạt động kết hợp của ít nhất 3 loại enzym endoglucanase (EC 3.2.1.4), exoglucanase (EC 3.2.1.74) và β-glucosidase (EC3.2.1.21) [52] (Hình 1.10).
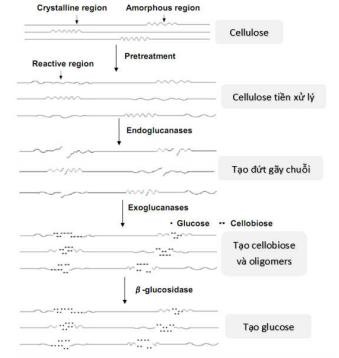
Hình 1.10 Hoạt động của các enzym cellulose [51]]
Sản phẩm phức hợp nhiều enzym hay thiết kế cellulosome là một trong những phương pháp được nghiên cứu gần đây để tối ưu hóa quá trình thủy phân sinh khối. Thiết kế cellulosome bao gồm các vật liệu với CBM và rất nhiều các modul kết dính từ các hệ cellulase khác nhau được sinh tổng hợp từ nhiều loài vi sinh vật. Các enzym từ các nguồn khác nhau được kết dính với nhau và thường được liên kết với một giá thể để tạo ra một phức hợp đa chức năng có kích thước lớn, gọi là cellulosome. Phức hợp cellulosome được cấu tạo bởi sự tương tác không cộng hóa trị và các liên kết đặc biệt, có ái lực cao giữa các mô-đun gắn kết. Trong trường hợp này, CBM liên kết với các thành phần trong cellulosome [50].
So với các hệ thống enzym đơn chức năng, phức hợp cellulosome thể hiện sức mạnh tổng hợp giữa các phân tử một cách chặt chẽ. Khi đưa các đơn vị cấu thành xúc tác có chức năng bất định vào liên kết vật lý chặt chẽ với nhau, hai enzym
có thể được coi là riêng biệt và có sức mạnh tổng hợp giữa các phân tử. Cả hệ thống enzym đơn chức năng và hệ thống cellulosome đều sử dụng các enzym họ celulase và hemicellulase. Sự tương tác tương hỗ giữa các enzym tạo nên một hệ phức lớn xúc tác linh hoạt,bổ sung lẫn nhau làm tăng hiệu quả quá trình thủy phân [50].
Mô hình mới thứ ba có thể được xem là trung gian giữa mô hình các enzym đơn chức năng và các enzym tổng hợp cao, đó là các enzym đa chức năng. Enzym đa chức năng là sản phẩm sinh tổng hợp từ một vi sinh vật nào đó, bao gồm hai hoặc nhiều enzym có mặt và cùng hoạt động xúc tác. Các enzym này thường bao gồm khối lượng phân tử cao và có một hoặc một số CBM. Trong tự nhiên, các enzym đa chức năng tồn tại trong cả hệ thống enzym đơn chức năng và hệ thống mô tế bào. Việc phân loại các enzym đa chức năng dựa trên các đặc tính cơ chất và do đó có thể được nhóm thành một trong bốn lớp khác nhau: cellulase –– cellulase, cellulase – hemicellulase, hemicellulase – hemicellulase, và hệ thống hemicellulase
– carbohydrate esterase [53].
1.3.4. Ứng dụng cellulase
Cellulase được ứng dụng nhiều trong các lĩnh vực khác nhau như trong sản xuất thức ăn gia súc, trong sản xuất thực phẩm và đồ uống, trong dệt may, trong sản xuất giấy và đặc biệt trong sản xuất nhiên liệu cồn sinh học [54].
Trong sản xuất bia, rượu và các loại đồ uống lên men từ ngũ cốc, cellulase chủ yếu hỗ trợ quá trình phân giải celluloase làm tăng hiệu quả quá trình lọc dịch đường sau thủy phân hoặc làm trong dịch lên men, giảm độ nhớt v.v…Các quá trình này chủ yếu sử dụng các cellulase dưới dạng các thương phẩm [55].
Trong chế biến thức ăn gia súc cellulase phân hủy chất xơ có trong nguyên liệu nhằm tăng giá trị chuyển hóa của thức ăn đối với động vật [56][57].
Trong công nghiệp dệt may cellulase được sử dụng với mục đích làm mềm, mịn, sạch và ổn định màu sắc của vải, trong các công đoạn của quá trình sản xuất endoglucanse được bổ sung để loại bỏ các sợi xơ ngắn trên bề mặt vải và các bụi bẩn trong mạng lưới vi sợi. Ngoài ra, trong sản xuất quần jean cellulase còn được sử dụng để tạo ra các vêt xước, rách “stonewashing” theo ý muốn mà không gây tổn hại đến vải, với cách này đã thay thế được [56].
Nhiên liệu sinh học là nguồn nguyên liệu thay thế các nguyên liệu hóa thạch quan trọng, ngày nay nhiều quốc gia đang tìm cách để giảm sự phụ thuộc vào nhiên liệu hóa thạch. Một trong những bước quan trọng trong sản xuất nhiên liệu sinh học là chuyển hóa lignocelluloses thành đường có thể lên men. Hiệu quả của quá trình này phụ thuộc vào mức độ phân giải hiệu quả của hệ enzym phân giải lignocellulose [58].
Sinh khối Lignocellulose là nguồn năng lượng tái tạo với trữ lượng lớn trên trái đất. Tuy nhiêu quy trình sản xuất ethanol sinh học từ sinh khối phức tạp hơn nhiều so với việc sản xuất ethanol từ tinh bột hay từ đường. Để giảm được chi phí cho quá trình này cần có nguyên liệu thô chi phí thấp, hệ enzym hiệu quả và các phương pháp tiền xử lý. Sinh khối cellulose là nguyên liệu có giá thành thấp, có thể tận dụng từ các phụ phẩm của các ngành khác và có thể tái tạo. Những vật liệu này bao gồm dăm gỗ, rơm rạ, bã mía, cây trồng, cỏ, lõi ngô v.v…Trong quá trình thủy phân đường đơn được tạo ra nhờ sự chyển hóa của hệ enzym cellulase và hemicellulase [24]
Quá trình thủy phân các bã thải nông nghiệp: bã mía, lõi ngô,...bằng enzym được nghiên cứu rộng rãi tùy thuộc vào mục đích và khả năng ứng dụng của các sản phẩm thu được sau khi thủy phân. De Guilherme và cs (2017) đã sử dụng một nhóm các enzym: phức hợp cellulase với β-glucosidase, phức hợp (arabinase, β-glucanase, cellulases, hemicellulases, pectinase, xylanase), β-glucosidase và xylanase để thủy phân bã mía được tiền xử lý bằng acid kết hợp với kiềm để tăng cường sản xuất glucose và xylose. Kết quả của nghiên cứu này cho thấy rằng quá trình kết hợp tiền xử lý này làm tăng lượng cellulose lên 68,3%, giảm 16,2% lượng hemicellulose do quá trình tiền xử lý trước bằng acid làm một lượng hemicellulose bị hòa tan. Lượng lignin trong sinh khối giảm 10,9% [59].
Sự kết hợp các enzym khác nhau để thủy phân làm tăng hiệu suất chuyển hóa bã mía thành đường, cải thiện nồng độ cuối cùng của glucose và xylose trong môi trường lên men. Việc sử dụng phức hợp có chứa xylanase trong hỗn hợp enzym đã làm tăng lượng glucose và xylose lên 12,7% và 378,4% so với các nghiên cứu trước [59].
Nhiều nghiên cứu đã sử dụng cellulase thu nhận từ vi sinh vật kết hợp các phương pháp tiền xử lý để thủy phân hiệu quả lignocellulose. Sulaiman và cộng sự sử dụng cellulase từ Geobacillus stearothermophilus để thủy phân rác thải giàu cellulose từ cây chà là đã được tiền xử lý, kết quả hiệu suất đường hóa đạt 71,03% [55]. Han và cộng sự cũng đã kết hợp phương pháp tiền xử lý bằng kiềm khi sử dụng cellulase thu nhận từ Penicillium waksmanii F10-2 để thủy phân rơm từ lúa mỳ, quá trình thủy phân đạt tối ưu ở điều kiện thời gian phản ứng 30 giờ, nồng độ cơ chất 3%, nhiệt độ 55ºC và pH 5,0 hàm lượng đường khử tạo thành cao nhất đạt 350mg/g cơ chất [60]. Tsai và cộng sự cũng đã kết hợp enzym thủy phân cellulose với các phương pháp tiền xử lý khác nhau khi thủy phân cỏ voi, kết quả nghiên cứu đã lưa chọn phương pháp tiền xử lý bằng NaOH với cellulase (CTec2) cho hiệu suất thủy phân cao nhất đạt 74,5% [61].
Radhika và cộng sự đã tối ưu hóa quá trình thủy phân sinh khối cây cao lương đã tiền xử lý bằng axit sử dụng cellulase từ Aspergillus sp, hiệu suất thủy phân cao nhất là đạt 55,97% [62].
Nhiều nghiên cứu đã chứng minh một số nhiên liệu sinh học như ethanol sinh học, khí sinh học … và một số hợp chất, dược phẩm quan trong trọng có thể được sản xuất từ nguồn lignocellulose. Sản xuất năng lượng sinh học từ nguồn lignocellulose thải được gọi là năng lượng sinh học thế hệ thứ hai. Khác với thế hệ thứ nhất là, năng lượng sinh học được sản xuất từ lương thực như bắp, sắn, đậu hay một số loại cây dầu, sự sản xuất năng lượng sinh học thứ hai không gây áp lực lên nhu cầu cung cấp lương thực. Trong đó sản xuất ethnol sinh học từ sinh khối lignocellulose thải nhận được nhiều sự quan tâm vì nó giải quyết vấn đề ô nhiễm môi trường và tăng lợi ích kinh tế [63]
Tại Việt Nam Phan Thị Phẩm và cộng sự đã nhận thấy tiềm năng trong việc chuyển đổi sinh khối lignocellulose trong sản xuất ethanol sinh học, tác giả đã nhận thấy các nguyên liệu lignocellulose rất dồi dào ở Việt Nam, ước tính 113,7 triệu tấn/năm và bằng cách tiếp cận với công nghệ mới để đẩy mạnh việc nghiên cứu sâu hơn để có thể áp dụng sản xuất ethanol sinh học từ nguồn lignocellulose thải [51].
1.4. Ứng dụng giải trình tự genome để phát hiện tiềm năng vi khuẩn sinh tổng hợp cellulase trong phân giải lignocellulose
Giải trình tự toàn bộ gen là phương pháp toàn diện nhất để phân tích dự đoán chức năng các gen liên quan [64]. Phương pháp giải trình tự toàn bộ gen đã được sử dụng trong nhiều nghiên cứu để thúc đẩy nhanh chóng trong việc khám phá ra các chủng sinh cellulase và hemicellulase tiềm năng.
Pasari và cộng sự giải trình tự hệ gen chủng vi khuẩn Paenibacillus polymyxa A18 được phân lập từ ruột mối, kết quả xác định được 4608 trình tự gen mã hóa, nhiều gen mã hóa các enzym liên quan đến thủy phân cacbohydrat, bao gồm các enzym chịu trách nhiệm thủy phân gỗ [65].
Nghiên cứu khác của Zheng và cộng sự bằng phương pháp giải trình tự bộ gen của chủng Cellulosimicrobium sp. TH-20 đã khám phá được các gen chức năng liên quan đến sự biến đổi ginsenoside. Ginsenoside là hoạt chất sinh học chính của nhân sâm giúp chống mệt mỏi, chống dị ứng, chống viêm, chống ung thư. Glycosidase được coi là công cụ hữu ích để xúc tác quá trình thủy phân các phân tử ginsenoit tạo ra ginsenoside. Chi Cellulosimicrobium đã thu hút được sự chú ý nhờ khả năng sinh glycosidase, bao gồm endo-beta-1,4-glucanase, endo-beta-1,4- xylanase, endo-beta-1,4-mannanase và endo-beta-1,3-glucanase [66]
Phân tích trình tự hệ gen vi khuẩn chịu mặn Parvularcula flava NH6-79T đã dự đoán 3465 gen, 96 gen trong số đó thuộc glycoside hydrolase (GHs). Trong các GH này, 20 gen được mã hóa có liên quan đến sự phân hủy cellulose, bao gồm 12 endoglucanase (5 GH10, 4 GH5 và 3 GH51), 2 exoglucanase (GH9) và 6 β- glucosidase (GH3). Ngoài ra, hoạt độ enzym tương đối cao nhất (endoglucanase, exoglucanase và β-glucosidase) được quan sát thấy ở giờ thứ 27 khi chủng được nuôi cấy trong môi trường chứa carboxymethyl cellulose/Avicel trong 45 giờ. Sự kết hợp giữa phân tích bộ gen với các nghiên cứu thực nghiệm đã chỉ ra khả năng của chủng NH6-79 T trong việc sinh tổng hợp endoglucanase, exoglucanase và β- glucosidase ngoại bào. Những phát hiện này cho thấy tiềm năng của chủng Parvularcula flava NH6-79 T trong việc phân hủy sinh khối chứa cellulose và chủng này có thể được sử dụng trong quá trình thu nhận cellulase [67].
Micromonospora sp CP22 được báo cáo là chủng có khả năng phân giải sinh khối lignocelluloses dựa trên 63 CAZyme được tìm thấy trong bộ gen đã được giải trình tự của chủng, theo công bố của An và cộng sự 2021 [68].
Bộ gen B. velezensis LC1 được chứng minh có 31 gen liên quan đến sự phân giải lignocellulose, trong đó còn có sự có mặt của các gen hỗ trợ, phát hiện này có ý nghĩa rất lớn trong việc đánh giá về hệ gen từ vi khuẩn sống cộng sinh trong ruột mối theo công bố của Li và cộng sự 2020 [69]
Nhằm nghiên cứu sự kết hợp của vật chủ mối và vi sinh vật đường ruột của chúng trong quá trình tiêu hóa lignocellulose, Tartar và cộng sự đã phân tích song song trình tự 2 thư viện cDNA của các sinh vật cộng sinh trong ruột mối và của vật chủ mối bậc thấp R. flavipes để tìm gen mã hóa lignocellulase của mối cũng như của vi sinh vật cộng sinh trong đường ruột của chúng. Cả 2 thư viện có tất cả 10.000 EST và dự đoán được chức năng của 6.555 EST. Trong số các EST dự đoán được, có 171 gen lignocellulase gồm 77 gen cellulase, 45 gen hemicellulase, 14 gen chitinase và 17 gen α-carbonhydrolase. Tỷ lệ các gen cellulase và hemicellulase của sinh vật ruột mối cao hơn nhiều so với của vật chủ mối. Cellulase của động vật nguyên sinh chiếm 66%, của sinh vật nhân sơ chiếm 18% trong tổng số các gen cellulase; gen hemicellulase của động vật nguyên sinh chiếm 69% và còn lại là của sinh vật nhân sơ. Ngoài ra, có 2 gen mã hóa dokerin và 7 gen mã hóa CBM (carbohydrate binding module) cellulase và hemicellulase cũng được dự đoán thuộc sinh vật nhân sơ đường ruột mối [70].