Bảng 1.2 Tính bền nhiệt của cellulase và xylanase từ C.cellulans [18].
Họat độ cellulse, xylanase tương đối còn lại (%) | |||
30 ºC | 40 ºC | 50ºC | |
1 giờ | 94,91 | 64,87 | 25,13 |
6 giờ | 80,92 | 55,68 | 10,13 |
Có thể bạn quan tâm!
-

 Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 1
Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 1 -
 Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 2
Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 2 -
![Đặc Tính Của Cellulose Và Xylanase Từ C.cellulans [18]](https://tailieuthamkhao.com/uploads/2023/02/19/nghien-cuu-cellulase-tu-vi-khuan-ruot-moi-phan-lap-o-viet-nam-3-1-120x90.jpg) Đặc Tính Của Cellulose Và Xylanase Từ C.cellulans [18]
Đặc Tính Của Cellulose Và Xylanase Từ C.cellulans [18] -
 Ứng Dụng Giải Trình Tự Genome Để Phát Hiện Tiềm Năng Vi Khuẩn Sinh Tổng Hợp Cellulase Trong Phân Giải Lignocellulose
Ứng Dụng Giải Trình Tự Genome Để Phát Hiện Tiềm Năng Vi Khuẩn Sinh Tổng Hợp Cellulase Trong Phân Giải Lignocellulose -
 Phương Pháp Phân Lập Và Tuyển Chọn Chủng Sinh Tổng Hợp Cellulase
Phương Pháp Phân Lập Và Tuyển Chọn Chủng Sinh Tổng Hợp Cellulase -
 Khảo Sát Khả Năng Thủy Phân Rơm Sử Dụng Cellulase Thu Nhận Từ Vi Khuẩn
Khảo Sát Khả Năng Thủy Phân Rơm Sử Dụng Cellulase Thu Nhận Từ Vi Khuẩn
Xem toàn bộ 138 trang tài liệu này.
Bảng 1.3 Tính bền pH của cellulase và xylanase từ C.cellulans [18].
Họat độ cellulse, xylanase tương đối còn lại (%) | |||
pH 3,4 | pH 5,0 | pH 7,0 | |
1 giờ | 22,22 | 50,65 | 40,46 |
6 giờ | 18,18 | 34,39 | 26,26 |
Yếu tố pH ngoài ảnh hưởng đến sinh trưởng và sinh tổng hợp enzym của vi sinh vật, nhiều nghiên cứu đã chỉ ra pH kiềm yếu có vai trò nhất định trong xử lý lignocellulose. Cụ thể, pH kiềm giúp nới lỏng thành tế bào của nguyên liệu lignocellulose, làm tăng phần diện tích bề mặt bên trong mà enzyme có khả năng tiếp cận, giảm sự polyme hóa, tinh thể hóa và làm tách các liên kết giữa lignin với phần carbohydrat bên trong. Do đó, điều kiện pH kiềm yếu có thể cải thiện hiệu quả quá trình phân hủy sinh khối lignocellulose [12].
1.1.4.2. Ảnh hưởng của các ion kim loại và các chất phụ gia đến hoạt độ enzym.
Cellulase bị ức chế bởi các sản phẩm phản ứng của nó như glucose, cellobiose và bị ức chế hoàn toàn bởi Hg. Ngoài ra cellulase còn bị ức chế bởi các ion kim loại khác như Mn, Ag, Zn nhưng ở mức độ nhẹ. Trọng lượng của enzym cellulase thay đổi từ 30-110 KDa (beguin, 1990) [28].
Các ion kim loại có thể kìm hãm hoặc hoạt hóa sự hoạt động của các enzym. Các ion kim loại nặng ở nồng độ nhất định có thể gây biến tính và kìm hãm không thuận nghịch enzym. Theo Sharma và cộng sự nhận thấy, ion Ca2+ làm tăng hoạt tính của cellulase từ Bacillus sp lên 40% so với đối chứng, còn Mg2+ làm giảm nhẹ (hoạt tính còn lại 92%) và Zn2+ ức chế hoạt tính của enzym này (hoạt tính còn lại bằng 37% so với đối chứng [22]. Với báo cáo của Nikky và cộng sự 201, endoglucanase từ Pseudomonas bị kìm hãm khi có mặt Zn2+, Cu2+ và Fe2+ trong khi Mn2+ làm tăng hoạt độ enzym [22].
Ngoài ra các dung môi hữu cơ, các chất tẩy rửa cũng ảnh hưởng mạnh mẽ đến hoạt tính của enzym. Tùy thuộc vào bản chất của các chất trên cũng như bản chất của enzym mà tính chất và mức độ ảnh hưởng tới hoạt động của enzym là khác
nhau. Theo nghiên cứu cho thấy, enzym bị giảm hoạt tính khi có mặt các chất tẩy rửa thông thường như Triton X100 và SDS ở nồng độ 5mM, ở nồng độ 1mM trong 1giờ thì nồng độ enzym còn lại khoảng 50%. Tuy nhiên, sự có mặt của các chất bề mặt không ion Tween 20 hoạt độ enzym tăng [22].
1.2. Tổng quan về Mối
Mối là tên gọi chung cho những côn trùng được xếp vào thứ bộ cánh đều (Isoptera) thuộc vào bộ Gián (Blattodea) nhưng có nhiều tác giả có thể xếp mối vào một họ lớn Termitoidea [29].
1.2.1. Phân loại Mối
Mối bao gồm hơn 2600 loài được chia thành 281 giống và hình thành nên 7 họ. Trong 7 họ này, chỉ có duy nhất họ Termitidae được xem là mối bậc cao với hơn 200 loài sống ở các môi trường sống khác nhau trên toàn thế giới. Dù thuộc loài nào thì các quần thể mối vẫn được phân thành các tầng lớp. Trong đó, mối thợ là những cá thể đảm nhận rất nhiều việc: tìm đồ ăn, nước, xây dựng, sửa chữa hệ thống đường hầm, chăm sóc cho các cá thể chưa trưởng thanh. Vì lý do này, mối thợ là đối tượng nghiên cứu chủ yếu của các nghiên cứu về hệ vi sinh vật thủy phân cellulose trong ruột mối [30].
Các loài mối khác nhau, chúng khác nhau về cấu trúc tổ (có loài làm tổ nổi trên mặt đất, có loài làm tổ chìm, có loài làm tổ trên cây), đặc điểm dinh dưỡng (có loài chuyên ăn gỗ khô, có loài chuyên ăn gỗ ẩm, có loài chuyên ăn mùn), có loài đắp đường mui, có loài không đắp đường mui khi đi kiếm ăn, có loài ăn bên ngoài có loài chuyên ăn bên trong gỗ [31] [32]. Ở Việt Nam, hiện đã phát hiện 106 loài mối. Trong đó có một số nhóm loài gây hại thường gặp là các giống: Coptotermes, Odototermes, Macrotermes, Microtermes, Hypotermes, Cryptotermes. Với các đối tượng nhà cửa, kho tàng; đê đập và cây... bị các loài mối gây hại ở mức độ khác nhau. Riêng với nhà cửa, kho tàng ở nước ta thì mức độ gây hại nghiêm trọng nhất thuộc về các loài mối thuộc giống Coptotermes (còn gọi là mối gỗ ẩm), tiếp theo là các loài thuộc nhóm mối đất, tập trung trong 2 giống Odontotermes và Macrotermes, cuối cùng là nhóm mối gỗ khô, thuộc giống Cryptotermes. Các loài mối Coptotermes làm tổ ngầm trong nền móng công trình, trong cây, hoặc kết cấu khác của công trình, đường mui chủ yếu là đơn lẻ, chúng hoạt động ở nhiều tầng cao thấp khác nhau của công trình.
Các loài mối đất chỉ làm tổ trong đất, trong tổ luôn có vườn nấm Termitomyces. Mối sống trong đất: Chúng thường dựa vào đất để làm tổ, thường ở gần phần rễ của cây hoặc trong cột gỗ chôn trong đất, tổ của nhóm mối này thường chìm trong đất hoặc nửa nổi nửa chìm trong đất
Các loài mối gỗ khô chỉ làm tổ trong các cấu kiện gỗ, số lượng cá thể của một tổ thường chỉ có vài trăm con, phân thải ra có dạng hạt cải. Loại tổ mối này thường làm trong gỗ hoặc trong cành cây khô không liên hệ với đất [33].
Thức ăn của mối chủ yếu có nguồn gốc từ thực vật hoặc các loại nấm được cấy trong tổ. Quá trình dinh dưỡng của mối diễn ra như sau: Thức ăn do mối thợ nuốt vào trong cơ thể sau đó mối thợ đem thể dịch thức ăn đã được tiêu hoá hoặc tiêu hoá một phần trong cơ thể đưa ra đường miệng hoặc bài tiết ra đường hậu môn để bón cho mối vua, mối lính, mối non. Quá trình tiêu hoá thức ăn diễn ra như vậy là nhờ trong ruột mối thợ có những vi sinh vật cộng sinh và những vi sinh vật này có khả năng phân huỷ cellulose thành các đường đơn làm nguồn dinh dưỡng cho chúng [34].
Năm 1856, Lespes là người đầu tiên miêu tả sự tồn tại của vi sinh vật trong ruột mối. Tuy nhiên, việc nghiên cứu mối quan hệ cộng sinh giữa mối và vi sinh vật chỉ được bắt đầu từ đầu thế kỷ 20. Năm 1923, một thí nghiệm nổi tiếng của Cleveland đã chứng minh được mối không thể tồn tại trên gỗ nếu không có sự hỗ trợ của vi sinh vật [35]. Chất kháng sinh làm thay đổi khu hệ sinh thái của vi khuẩn trong ruột mối, phá vỡ sự tương tác cộng sinh giữa vi khuẩn và vật chủ, qua đó dẫn tới chu kỳ sống và khả năng sinh sản của mối bị giảm [35]
Mối là một loại côn trùng được biết đến với sức phá hủy lớn đối với các công trình xây dựng, đồ đạc vì chúng là đối tượng chính tác động đến những vật liệu được làm từ gỗ. Nhưng cũng chính vì vậy, chúng đã thu hút sự chú ý của các nhà nghiên cứu về việc phân hủy gỗ và lignocellulose. Có rất nhiều sinh vật trong ruột mối bao gồm vi sinh vật và nguyên sinh vật và tất cả đều là cần thiết cho quá trình tiêu hóa thức ăn của mối [30]. Vì lý do này mà hệ vi sinh vật cộng sinh trong ruột mối rất được chú ý nghiên cứu về đặc tính, cũng như các khả năng sinh enzym của chúng [31].
1.2.2. Hệ vi sinh vật trong ruột mối
Các loài mối phụ thuộc rất nhiều vào các enzym do vi sinh vật tạo ra để có thể phân hủy được cellulose. Có những loại mối mà việc thủy phân do hệ vi sinh vật cộng sinh trong ruột có thể chiếm đến hơn 70% quá trình thủy phân trong các đoạn ruột [36].
Quần xã vi sinh vật trong ruột mối có sự đa dạng rất cao. Nhiều nghiên cứu về hệ vi khuẩn trong ruột mối được thực hiện thông qua các phương pháp truyền thống dựa trên việc nuôi cấy và phân lập vi sinh vật ở trong phòng thí nghiệm. Các vi khuẩn Staphylococcus và Bacillus là những vi khuẩn có mặt nhiều nhất trong cả
ruột mối bậc thấp và bậc cao. Serratia marcescenes, Enterobacter aerogenes, Enterobacter cloacae và Citrobacter farmeri đã được phân lập từ Coptotermes formosanus trong khi đó E. aerogenes, E. cloacae và Clavibacter agropyri đã được chứng minh là tồn tại trong ruột mối C. curvignathus. Tương tự, Husseneder và cộng sự cũng đã phân lập từ C. formosanus 25 chủng vi khuẩn, thuộc các họ Enterobacteriaceae, bộ Bacteroidales và Lactobacillales [37].
Hongoh và cộng sự (2006) đã xác định được vi khuẩn ngành Fibrobacteres (chiếm khoảng 10%) cùng với vi khuẩn thuộc bộ Bacteroidales và Clostridiales chỉ đứng sau chi Treponema về độ đông đúc trong đường ruột của mối Microcerotermes Spirochaetes [38].
Gần đây, các nghiên cứu không thông qua nuôi cấy đã chứng minh nhiều hơn về sự đa dạng vi sinh vật ruột mối, đặc biệt là đa dạng vi khuẩn. Bằng phân tích trình tự 16S rRNA của vi khuẩn được khuếch đại từ metagenome của hệ vi sinh ruột mối, tổng số hơn 1500 phylotype (tương đương với loài, tuy nhiên không nuôi cấy và định tên được, chỉ có thể xác định được thông qua trình tự 16S rRNA) của vi khuẩn đã được phân tích và được phân loại vào 24 nhóm ngành khác nhau. Phần lớn chúng chưa bao giờ được tìm thấy ở trong các môi trường sống khác và cho thấy độ tương đồng thấp với các chủng vi khuẩn nuôi cấy được [17].
Ở Việt Nam đã có các nghiên cứu tiếp cận phương pháp metagenomic để đánh giá về sự đa dạng của hệ vi sinh vật trong ruột mối. Với phương pháp metagenomic (nghiên cứu đa bộ gen) có sự nổi trội vượt bậc vì có thể đánh giá được sự đa dạng về gen trong một quần thể vi sinh vật và đặc biệt nhất đó là có thể cho thấy được cả sự tồn tại của cả những cá thể không thể nuôi cấy được. Và bằng phương pháp này, hệ vi sinh vật trong ruột mối thuộc loài Coptotermes gestroi đã được phân tích và cho ra kết quả trong hình 1.1
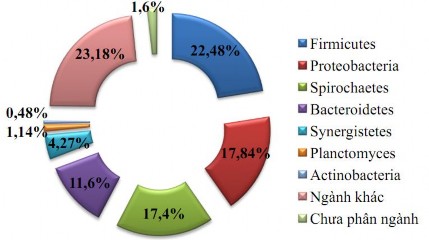
Hình 1.3. Hệ vi sinh vật trong ruột mối [39]
Chi mối Coptotermes được nghiên cứu Nguyễn Thanh Ngọc và cộng sự là chi mối có mặt phổ biến nhất tại Việt Nam, và sự đa dạng sinh học trong ruột mối có thể thấy, Firmicutes, Proteobacteria, Spriochaetes là 3 ngành vi khuẩn chiếm số lượng lớn nhất trong ruột mối này nhưng vẫn có một lượng lớn những ngành vi khuẩn khác và ngoài ra thì còn sự có mặt của những vi khuẩn chưa được phân ngành. Xạ khuẩn Actinobacteria cũng có mặt trong ruột mối nhưng với một số lượng vô cùng khiêm tốn so với các ngành khác [17].
Các nghiên cứu gần đây của Đỗ Thị Huyền, Nguyễn Thị Thảo và cộng sự cũng đã sử dụng phương pháp giải trình tự toàn bộ hệ gen vi khuẩn trong ruột mối để đánh giá sự đa dạng vi khuẩn có khả năng sinh cellulase từ ruột mối ở Việt Nam.
1.2.3 Phân lập vi khuẩn sinh cellulase từ ruột mối
Có nhiều phương pháp để nghiên cứu hệ enzym trong ruột mối, đó là các phương pháp tách các enzym từ ruột mối và nghiên cứu các đặc tính của chúng, phương pháp khác là phân lập, nuôi cấy các vi sinh vật trên môi trường tổng hợp một cách riêng rẽ sau đó tìm ra được các chủng có khả năng sinh enzym và thực hiện các phương pháp lên men để thu nhận. Với phương pháp phân lập chỉ có rất ít vi sinh vật có thể nuôi cấy được mới có thể sử dụng. Phương pháp mới gần đây nữa là các phương pháp sinh học phân tử nhằm tìm kiếm các gen có khả năng mã hóa sinh tổng hợp các enzym từ đó đã phát hiện ra một số vi sinh vật tiềm năng trong ruột mối
Rất nhiều nghiên cứu đã tìm cách phân lập vi khuẩn sinh cellulase từ ruột mối với mong muốn tìm được vi khuẩn có khả năng sinh cellulase cao hay hệ cellulase thủy phân hiệu quả cellulose. Theo Wenzel và cộng sự, lượng vi khuẩn sinh cellulase trong ruột mối vào khoảng 107TB/ml ruột mối nghiền [9]. Paenibacillus lactis, Lysinibacillus macrolides, Stenotrophomonas malttophilia,
Lysinibacillus fusifimics và Bacillus cereus là các loài vi khuẩn phân lập được từ mối đất Psammotermes hypostoma Desneux thuộc họ Rhinotermitidae ở Ai Cập
[40] và đều có hoạt tính endoglucanase dao động từ 0,22 U/ml đến 2,28 U/ml. Trong số các chủng vi khuẩn phân lập được từ ruột mối ăn gỗ tại Ấn Độ có
70% thuộc họ Bacillaceae, 23% thuộc họ Enterobacteriaceae, 7% thuộc họ Paenibacillaceae. Các chủng đều thể hiện hoạt tính endoglucanase thêm 1 hay nhiều hơn hoạt tính cellulase như cellobiohydrolase hoặc avicelase hoặc β-glucosidase. Ngoài ra nhiều chủng còn biểu hiện hoạt tính hemicellulase như mannase, xylanase [41]. Trong nghiên cứu của Ferbiyanto, 2 chủng phân lập được từ mối Macrotermes gilvus sp thuộc họ Termitidae thu thập tại Indonesia có tỷ lệ đường kính vòng thủy phân/đường kính khuẩn lạc cao từ 2,81-3,5 các chủng này đã được định tên kết quả cho thấy chúng thuộc loài Bacillus megaterium và Paracoccus yeei
[42].
Ba chủng vi khuẩn thuộc các chi Acinetobacter, Pseudomonas, Staphylococus và bốn chủng thuộc họ Enterobacteriaceae, Bacillaceae được phân lập từ mối Microcerotermes diversus thuộc họ Termitidae thu thập tại Iran. Trong đó 2 chủng Bacillus sp B5B và Acinetobacter sp.L9B có hoạt tính endoglucanase cao nhất đạt 1,47 U/ml và 1,22 U/ml tương ứng [43]. Nghiên cứu của Wenzel và cộng sự đã phân lập được 119 chủng vi khuẩn có khả năng sinh cellulase từ mối bậc thấp Zootermopsis angusticoliss. Các chủng phân lập được thuộc các chi Cellulomonas/Oerskovia, Microbacterium, Kocuria, Bacillus, Brevibacillus, Paenibacillus, Afipia, Agrobacterium/Rhizobium, Brucella/Ochrobactrum, Pseudomonas và Sphingomonas/Zymomonas trong đó phần lớn các chủng thuộc 2 chi Bacillus và Paenibacillus. Các chủng có vòng thủy phân lớn chủ yếu thuộc chi Cellulomonas, Bacillus và Paenibacillus [9].
Hiện tại có nhiều vi khuẩn trong ruột mối vẫn chưa nuôi cấy được do việc tạo điều kiện môi trường nuôi cấy tương tự như trong ruột mối là khó khăn. Không một loài nào được phân lập trong môi trường yếm khí (100% và85%/15%). Các loài được phân lập chủ yếu trong môi trường hiếu khí hoặc vi hiếu khí, do còn một phần oxy có thể thấm qua thành ruột và không thể ngăn cản sự truyền oxy trong quá trình nghiền thức ăn của ruột [31].
Khoảng 42 khuẩn lạc đã được lựa chọn theo phương pháp sàng lọc dựa trên phân tích sinh học và phân tích hoạt động enzyme. Phân tích trình tự 16S rDNA cho thấy tất cả các loài vi khuẩn đã xác định thuộc 5 họ - 50% thuộc về Bacillaceae, 26% thuộc về Enterobacteriaceae, 17% thuộc về Microbacteriaceae, 5% thuộc về
Paenibacillaceae và 2% thuộc về Promicromonosporaceae. Từ 60 đến 70% vi khuẩn từ ruột của mối đến từ Họ Bacillaceae (Hình 1.3) [41]
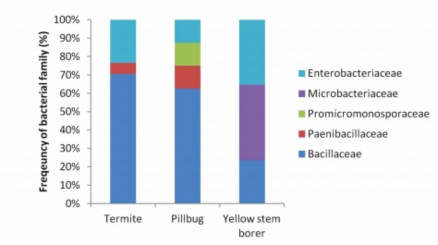
Hình 1.4 Phân bố các họ vi khuẩn trong ruột mối và côn trùng [41].
Bảng 1.4 liệt kê các loài vi khuẩn được phân lập từ các loài mối khác nhau cùng với khả năng sinh tổng hợp cellulase của chúng. Có thể thấy các chủng thuộc chi Bacillus chiếm lượng lớn các loài vi khuẩn phân lập được ở hầu hết các loài mối. Các vi khuẩn từ ruột mối có khả năng biểu hiện hầu hết hoạt tính của hệ cellulase như Endoglucanase, Cellobiohydrolase, -glucosidase, Filter cellulase tuy nhiên hoạt tính chiếm đa số là Endoglucanase
Bảng 1.4. Các vi khuẩn phân lập từ các loài mối [31].
Enzym | Loài mối | |
Acinetobacter sp. | EG, βG | Coptotermes curvignathus |
B. cereus | EG, FP | Zootermopsis angusticollis |
B. cereus | EG | Reticulitermes Hesperus |
B. cereus | EG, βG | Coptotermes curvignathus |
B. circulans | EG, FP | Zootermopsis angusticollis |
B. megaterium | EG, FP | Zootermopsis angusticollis |
Bacillus sp. | βG | Reticulitermes santonensis |
Bacillus sp. | CBH,βG,EG | Reticulitermes speratus |
Cellulomonas sp. | EG, FP | Zootermopsis angusticollis |
Chryseobacterium sp. | CBH, βG, EG | Reticulitermes speratus |
Citrobacter sp. | CBH, βG, EG | Reticulitermes speratus |
Clostridium beijerinckii | βG | Coptotermes formosanus |
Clostridium termitidis sp. | EG, βG | Nasutitermes lujae |
Comamonas sp. | CBH, βG, EG | Reticulitermes speratus |
CBH, βG, EG | Reticulitermes speratus | |
Enterobacter aerogenes | EG, βG | Coptotermes curvignathus |
Klebsiella sp. | βG | Reticulitermes santonensis |
Klebsiella sp. | CBH, βG, EG | Reticulitermes speratus |
Kocuria varians | EG, FP | Zootermopsis angusticollis |
Microbacterium sp. | EG, FP | Zootermopsis angusticollis |
Ochrobactrum sp. | EG, FP | Zootermopsis angusticollis |
Paenibacillus sp. | EG, FP | Zootermopsis angusticollis |
Paenibacillus sp. | EG, FP | Microcerotermes diversus |
Serratia marcescens | EG | Reticulitermes Hesperus |
Serratia sp. | CBH, βG, EG | Reticulitermes speratus |
Sphingomonas echinoides | EG, FP | Zootermopsis angusticollis |
Sphingomonas sp | EG, FP | Zootermopsis angusticollis |
Staphylococcus sp. | EG, FP | Microcerotermes diversus |
Streptomyces sp. | EG, FP | Armitermes |
Dyella sp.
EG : Endoglucanase activity – FP : Filter Paper-cellulase activity – βG : β- glucosidase activity – CBH : Cellobiohydrolase activity
1.3. Ứng dụng cellulase trong phân giải sinh khối
1.3.1. Thành phần Lignocellulose
Lignocellulose là thành phần chính của thực vật thân gỗ và các thực vật khác như cỏ, lúa, ngô…gồm 3 hợp phần chính là cellulose, hemicellulose và lignin
Cellulose
Cellulose là polyme đồng nhất có cấu trúc ổn định, là polyme mạch thẳng của các phân tử glucose liên kết với nhau bằng liên kết β-1,4 glucosid. Cellulose bao gồm cả phần tinh thể và phần vô định hình, từ các phân tử D-gluocose. Cellulose không giống như tinh bột không cuộn xoắn và không phân nhánh [44]. Dạng tồn tại khác đó là việc hình thành nên các dạng hình que hoặc dạng sợi bằng việc tạo ra các nhóm lân cận phân từ glucose. Nhiều nhóm OH trên phân tử glucose từ một chuỗi lên kết H với O tạo với các nhóm OH khác hình thành nên các chuỗi giống nhau, liên kết với nhau tạo nên hệ vi sợi vững chắc. Cellulose có thể hình thành nên các hệ sợi nhỏ là nhờ các liên kết bên trong hoặc bên ngoài chuỗi tạo nên cấu trúc cứng và ổn định của cellulose. Cấu trúc vững chắc này đóng vai trò quan trọng trong cấu tạo của thành tế bào, tại đó các hệ vi sợi là các mắt lưới hình thành nên các ma trận hydratcacbon, cấu tạo này liên quan tới sự vững chắc của tế bào