như lạnh giá hay khô hạn, chúng tồn tại trong đất, trên tàn dư thực vật hay các ký chủ phụ, cỏ dại, trong cơ thể của ốc sên vào mùa khô và là nguồn bệnh lây nhiễm cho cây trồng vụ sau. Khi mùa mưa đến, bào tử hậu và bào tử trứng nảy mầm hình thành nên cành mang bọc bào tử động và giải phóng bào tử động xâm nhiễm cây hồ tiêu [44].
1.3.2. Bệnh chết chậm trên cây hồ tiêu do nấm Fusarium sp. và tuyến trùng
Bệnh vàng lá chết chậm là một trong những loại bệnh phổ biến trên cây hồ tiêu (Piper nigrum L.). Nguyên nhân gây bệnh vàng lá có thể do nhiều tác nhân sinh học cũng như phi sinh học khác nhau bao gồm nấm Fusarium sp., Rhizoctonia sp.; tuyến trùng ký sinh (Radopholus similis và Meloidogyne incognita), cây thiếu dinh dưỡng, nhiệt độ hay bị stress do nước. Fusarium solani f. sp. piperis được xem là tác nhân gây bệnh thối rễ trên cây hồ tiêu trong khi đó. F. oxysporum Schl. f. sp. piperis ít phổ biến nhưng được xem là bệnh quan trọng trên cây hồ tiêu [45]. Để mô tả đặc tính của tác nhân gây bệnh vàng lá trên cây hồ tiêu ở Malaysia, 53 chủng Fusarium đã được phân lập từ rễ và từ đất vùng rễ cây hồ tiêu tại các vùng trồng tiêu chủ yếu ở Sarawak và Johor. Trong đó có 34 chủng được xác định là nấm F. solani và 19 chủng là nấm F. proliferatum dựa trên các đặc điểm hình thái và phương pháp giải trình tự gen tại vùng ITS1 và ITS2 của DNA ribosome 5,8S. Để xác định nguyên nhân gây bệnh vàng lá trên cây hồ tiêu, nghiên cứu đã tiến hành thử nghiệm về khả năng gây bệnh trên cây hồ tiêu 3 tháng tuổi, kết quả cho thấy, F. solani là nguyên nhân gây bệnh vàng lá trên cây hồ tiêu [46]. Theo Eng (2002) thì tuyến trùng Meloidogyne có quan hệ với nấm Fusarium. Tuyến trùng chích hút, tạo vết thương vùng rễ hồ tiêu và tạo cơ hội cho nấm Fusarium tấn công rễ, gây bệnh cho cây hồ tiêu. Năng suất hồ tiêu sẽ giảm nghiêm trọng nếu có sự kết hợp của hai tác nhân gây bệnh là tuyến trùng rễ Meloidogyne và nấm [47]. Nghiên cứu của Thuy và cộng sự (2013) đã phân lập được 09 loài nấm từ rễ cây hồ tiêu trong đó có sự hiện diện của nấm Fusarium solani. Nghiên cứu cũng xác định nguyên nhân chính gây bệnh vàng lá trên cây hồ tiêu không phải do nấm Fusarium solani mà do tác nhân khác đó là tuyến trùng M. incognita. Khi đánh giá tác động của
M. incognita riêng lẽ hoặc kết hợp với Fusarium solani trong điều kiện nhà lưới cho thấy cả hai đều làm tăng tỷ lệ cây hồ tiêu bị vàng lá và tác động đến sự sinh trưởng của cây. Đặc biệt vào thời điểm hai tuần sau khi lây nhiễm F. Solani, cây hồ tiêu không có biểu hiện nhưng lây nhiễm với M. incognita cây hồ tiêu có biểu hiện bệnh thông qua sự hình thành nốt sần trên rễ [8].
1. Nấm Fusarium
* Phân loại: Nấm Fusarium sp. thuộc giới Fungi; Ngành Ascomycota; Lớp Sordariomycetes; Bộ Hypocreales; Họ Nectriaceae; Chi Fusarium [48].
* Sự phân bố: Fusarium là loài nấm phổ biến, có mặt ở hầu hết các hệ sinh thái như trong đất, dưới lòng đất, không khí, các mảnh vỡ thực vật và các chất hữu cơ khác. Chúng phổ biến ở các vùng nhiệt đới, ôn đới và cũng có thể tìm thấy ở vùng sa mạc, núi cao và cực Bắc, nơi điều kiện khí hậu khắc nghiệt. Việc phân bố rộng rãi các loài Fusarium có thể do chúng có khả năng phát triển trên nhiều loại chất nền và khả năng phân tán trong tự nhiên. Chi Fusarium bao gồm nhiều tác nhân gây bệnh khác nhau. Chúng có khả năng biến đổi các đặc tính sinh học như tổng hợp các chất chuyển hóa thứ cấp đặc biệt là các độc tố gây hại thực vật, côn trùng và các vi sinh vật khác [48].
* Đặc điểm hình thái, cấu tạo
Chi Fusarium là một trong những chi nấm lớn nhất trên thế giới; bao gồm nhiều loài sản xuất độc tố gây bệnh cho nhiều loại cây trồng nông nghiệp quan trọng. Hệ sợi nấm Fusarium phân nhánh, có vách ngăn, sợi nấm thường không màu và chuyển màu nâu khi già. Hệ sợi nấm sản sinh độc tố tiết vào hệ mạch gây héo rũ cây chủ. Cơ thể dinh dưỡng dạng sợi đa bào, phân nhánh phức tạp, vách ngăn có lỗ thủng ở giữa, trong tế bào có một hoặc nhiều nhân, vách tế bào bằng chitin, glucan [49-50].
Nấm Fusarium phát triển nhanh trên môi trường PDA ở nhiệt độ 25ºC và hình thành tản nấm có hình thể tơi xốp như bông, bằng phẳng hoặc lan rộng trên môi trường nuôi cấy. Mặt trên của tản nấm có thể có màu trắng, kem, vàng, vàng cam, đỏ, tím hồng hoặc tím. Mặt dưới có thể không màu, vàng cam, màu đỏ, màu tím sẫm hay màu nâu [51]. Nghiên cứu sinh địa lý học chỉ ra rằng, các loài Fusarium khác nhau sẽ phân bố ở vùng địa lý khác nhau. Sự thay đổi trong cấu trúc và đa dạng nấm có thể do kết hợp với các vùng khí hậu đặc biệt trên thế giới [52].
Có thể bạn quan tâm!
-

 Tuyển chọn, nghiên cứu đặc tính kháng tác nhân gây bệnh và tạo chế phẩm phòng trừ bệnh rễ của các chủng vi khuẩn vùng rễ cây hồ tiêu Piper nigrum L. tại Tây Nguyên - 1
Tuyển chọn, nghiên cứu đặc tính kháng tác nhân gây bệnh và tạo chế phẩm phòng trừ bệnh rễ của các chủng vi khuẩn vùng rễ cây hồ tiêu Piper nigrum L. tại Tây Nguyên - 1 -
 Tuyển chọn, nghiên cứu đặc tính kháng tác nhân gây bệnh và tạo chế phẩm phòng trừ bệnh rễ của các chủng vi khuẩn vùng rễ cây hồ tiêu Piper nigrum L. tại Tây Nguyên - 2
Tuyển chọn, nghiên cứu đặc tính kháng tác nhân gây bệnh và tạo chế phẩm phòng trừ bệnh rễ của các chủng vi khuẩn vùng rễ cây hồ tiêu Piper nigrum L. tại Tây Nguyên - 2 -
 Thực Trạng Sản Xuất Hồ Tiêu Của Việt Nam Và Các Tỉnh Tây Nguyên
Thực Trạng Sản Xuất Hồ Tiêu Của Việt Nam Và Các Tỉnh Tây Nguyên -
 Các Giải Pháp Tổng Hợp Để Kiểm Soát Dịch Bệnh Trên Cây Hồ Tiêu Và Sản Xuất Bền Vững
Các Giải Pháp Tổng Hợp Để Kiểm Soát Dịch Bệnh Trên Cây Hồ Tiêu Và Sản Xuất Bền Vững -
 Tình Hình Nghiên Cứu Về Vi Sinh Vật Vùng Rễ Trên Cây Hồ Tiêu
Tình Hình Nghiên Cứu Về Vi Sinh Vật Vùng Rễ Trên Cây Hồ Tiêu -
 Phương Pháp Tuyển Chọn Các Chủng Vi Khuẩn Phân Lập Từ Vùng Rễ Cây Hồ Tiêu Tại Tây Nguyên, Định Danh Các Chủng Có Tiềm Năng Kháng Phytophthora, Fusarium
Phương Pháp Tuyển Chọn Các Chủng Vi Khuẩn Phân Lập Từ Vùng Rễ Cây Hồ Tiêu Tại Tây Nguyên, Định Danh Các Chủng Có Tiềm Năng Kháng Phytophthora, Fusarium
Xem toàn bộ 224 trang tài liệu này.
* Hình thức sinh sản của nấm Fusarium sp.: Fusarium có 2 hình thức sinh sản: Sinh sản sinh dưỡng và sinh sản vô tính. Do thiếu giai đoạn sinh sản hữu tính trong vòng đời nên gọi chung là nấm không hoàn chỉnh hay nấm bất toàn [40].
Sinh sản sinh dưỡng: Sợi nấm: từ một sợi nấm riêng rẽ, khi gặp điều kiện thuận lợi sẽ sinh trưởng và phân nhánh thành hệ sợi nấm. Bào tử hậu: là những tế bào hình tròn, có tế bào chất được cô đặc lại [53] có màng dày bao bọc [54], thỉnh thoảng
có bào tử hậu với vách tế bào xù xì hoặc có sắc tố (hình 1.2) [51]. Chất dinh dưỡng được chuyển từ tế bào kề bên sang tế bào ưu tiên làm tế bào này phồng lên trong một thời gian khá dài. Khi gặp điều kiện thuận lợi chúng sẽ nảy mầm và phát triển thành sợi nấm mới. Bào tử hậu có thể nằm ở giữa sợi nấm hoặc ở tận cùng của nó [53-54]; có thể ở dạng đơn lẻ, dạng cặp đôi, dạng chuỗi hay dạng cụm (hình 1.2). Bào tử hậu có thể quan sát được dưới kính hiển vi sau 10- 14 ngày. Nhưng nhiều loài Fusarium có thời gian hình thành bào tử chậm hơn (20- 42 ngày) và có khí khổng tạo ra bào tử. Một số loài Fusarium có tạo bào tử hậu như F. chlamydosporum, F. napiforme, F. oxyspoum, F. semitectum, F. solani, F.sporotrichoides, F.equiseti, F. tricinctum [51], [55].
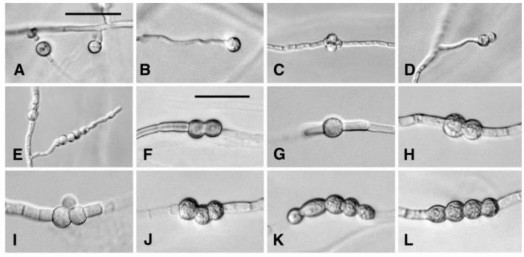
Hình 1.2. Bào tử hậu của các loài Fusarium [45], [50]
A-B: Bào tử hậu đơn, như hột cơm ở F. solani.; C-D: Bào tử hậu dạng chùm ở F. compactum; E: Bào tử hậu dạng chuỗi hột cơm ở F. compactum; F: Bào tử hậu dạng đôi có vách nhẵn ơ F. solani; G: Bào tử hậu đơn, dạng hột cơm ở F. scirpi; H: Bào tử hậu dạng đôi, dạng hột cơm ở F. compactum; I: Bào tử hậu dạng chùm, có vách nhẵn ở F. scirpi; J và L: Bào tử hậu dạng chuỗi hột ở F. compactum; K: Bào tử hậu dạng chuỗi hột ở F. scirpi. A-E: Tỷ lệ = 50 μm; F-L: Tỷ lệ = 25 μm.
Sinh sản vô tính: Bào tử đính được hình thành ở các loài nấm bất toàn. Đa số bào tử đính thường xếp thành chuỗi, có khi thành từng khối, một số bào tử đính nằm đơn độc từng cái trên một cuống bào tử đính. Cuống bào tử đính có thể đơn bào hoặc đa bào, không phân nhánh hoặc phân nhiều nhánh, mọc riêng lẻ hay sắp xếp từng cụm [56]. Ở các loài Fusarium thì bào tử đính thường là bào tử ngoại sinh, có 2 loại: bào tử đính lớn và bào tử đính nhỏ.
Bào tử đính lớn (bào tử lớn): trong suốt, được hình thành từ thể bình, trên cành
bào tử có nhánh hay không có nhánh [40]. Bào tử lớn có kích thước 2-8 x 11-70 µm. Kích thước thường dùng để phân biệt giữa các loài Fusarium: bào tử quá dài (F. avenaceum, F. coccophilum, F. decemellulare...) và quá ngắn (F. xylarioides, F. larvanrum, F. poae...). Phần đầu và phần cuối bào tử lớn có hình dạng và kích thước khác nhau. Phần đầu bào tử có thể ở dạng: cái móc, hình núm vú, hình đầu tù, hình nón. Phần cuối bào tử có dạng: hình đầu tù, hình núm vú, hình bàn chân, hình bàn chân được kéo dài, hình thon dài với vết khía hình chữ V (hình 1.3)[51], [55].
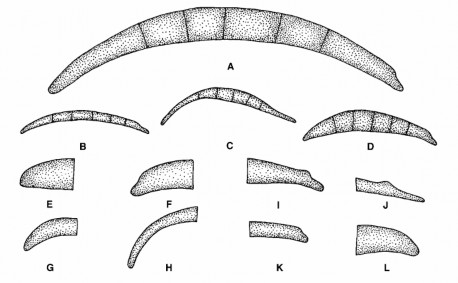
Hình 1.3. Bào tử đính lớn [57]
A-D: Hình dạng bào tử đính lớn: A. Bào tử đính lớn điển hình của Fusarium. Tế bào đỉnh bên trái, tế bào đáy bên phải. B. Bào tử đính dạng mảnh, thẳng và gần giống kim, e.g., F. avenaceum. C. Bào tử đính với độ cong mặt lưng, e.g., F. equiseti. D. Bào tử đính lớn có mặt lưng cong hơn mặt bụng, e.g., F. crookwellense.; E-H: Hình dạng tế bào đỉnh của bào tử đính lớn. E. Bằng, ví dụ, F. culmorum. F. có nhú, ví dụ, F. sambucinum. G. Có móc, ví dụ, F. lateritium. H. Thon, ví dụ, F. equiseti. I-L: Hình dạng tế bào đáy của bào tử đính lớn. I. Hình bàn chân, ví dụ, F. crookwellense. J. Hình dạng bàn chân kéo dài, ví dụ, F. longipes. K. Vết khía một cách riêng biệt, ví dụ, F. avenaceum. L. Vết khía vừa đủ, ví dụ, F. solani.
Bào tử đính nhỏ (bào tử nhỏ): kích thước 2-4 x 4-8 µm, được hình thành từ cành bào tử phân sinh phân nhánh hoặc không phân nhánh, mọc trực tiếp từ sợi nấm hoặc tụ lại thành dạng bọc giả trên đầu cành hoặc hình thành dạng chuỗi (hình 1.4) [58].
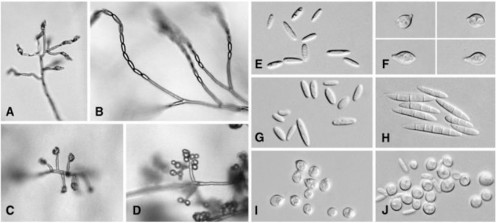
Hình 1.4. Sự hình thành và các loại bào tử đính nhỏ do các loài Fusarium tạo ra (Nguồn: Summerell và cộng sự, 2003 [59])
A: bào tử đính nhỏ sinh ra thành chuỗi ngắn (F. brevicatenulatum). B: bào tử đính nhỏ tạo thành chuỗi dài (F. Decemcellulare). C: bào tử đính nhỏ sinh ra ở đầu giả (F. Circinatum). D: bào tử đính nhỏ Napiform dạng ở đầu giả (F. konzum). E: bào tử đính nhỏ hình bầu dục (F. babinda). F: bào tử đính nhỏ Pyriform (F. anthophilum). G: bào tử đính nhỏ Clavate (F. anthophilum). H: bào tử đính nhỏ Fusiform (F. semitectum). I: bào tử đính nhỏ Napiform (F. poae). J: bào tử đính nhỏ hình cầu (F. anthophilum).
* Mức độ gây hại của nấm Fusarium sp.
Fusarium có thể tồn tại trong đất một thời gian dài mà không cần vật chủ, chủ yếu ở dạng bào tử áo có thành dày. Bào tử nảy mầm khi tiếp xúc gần với rễ và bắt đầu lây nhiễm, sợi nấm bám vào lớp vỏ rễ và xâm nhiễm tới các tế bào bên trong cho đến khi nó đến các mạch xylem và đi vào bên trong mạch thông qua các khoảng không gian giữa các tế bào. Tại đây, nấm chuyển sang giai đoạn lây nhiễm đặc biệt, chúng chỉ tồn tại chủ yếu trong các mạch xylem để nhanh chóng xâm chiếm vật chủ. Điều này chủ yếu được thực hiện bằng cách sản xuất các bào tử đính nhỏ, được tách ra và chuyển vào dòng nhựa của cây. Bào tử đính tiếp tục nảy mầm và sợi nấm xâm nhập vào phía bên trên thành mạch tạo ra nhiều bào tử đính ở các mạch tiếp theo. Triệu chứng héo bắt đầu xuất hiện do thiếu nước nghiêm trọng, chủ yếu là do tắc nghẽn mạch. Nguyên nhân héo có thể do sự kết hợp của nhiều hoạt động như sự tích tụ của sợi nấm, phản ứng phòng vệ của cây chủ hay sản xuất độc tố, bao gồm sản xuất gel, gum và tylose và sự ép vỡ mạch bởi sự tăng nhanh của các tế bào nhu mô liền kề. Khi cây còn sống, nấm chỉ có trong mô xylem và một số tế bào xung quanh. Chỉ khi cây nhiễm bệnh và chết, nấm mới xâm chiếm nhu mô và sinh sản trên bề mặt cây. Quá trình lây nhiễm của Fusarium diễn ra phức tạp với nhiều bước được kiểm soát một cách chặt chẽ (1) nhận biết rễ thông qua các các tín hiệu vật chủ không xác
định; (2) gắn kết lên bề mặt rễ và sự phân hóa của sợi nấm xâm nhập; (3) xâm nhập vào vỏ rễ và phá hủy cấu tạo rễ của vật chủ như lớp vỏ trong để tiếp cận mô mạch; (4) thích ứng với môi trường phòng vệ của vật chủ, bao gồm khả năng chống lại các hợp chất kháng nấm thực vật; (5) phát triển sợi nấm và sản xuất bào tử đính trong các mạch xylem và (6) tiết ra các độc lực quyết định như các peptide hoặc các phytotoxin [60]. Hầu hết các chủng Fusarium sp. đều có khả năng tổng hợp hàng loạt các chất chuyển hóa thứ cấp và các hợp chất này gây ảnh hưởng đến thực vật, động vật và sức khỏe con người.
2. Tuyến trùng
Tuyến trùng là nguyên nhân chính dẫn đến tình trạng suy thoái về chất dinh dưỡng và gây thiệt hại nghiêm trọng đối với hầu hết các loại cây trồng. Sau khi nở từ trứng, tuyến trùng non xâm nhập vào hệ thống rễ cây đang phát triển, sử dụng các chất dinh dưỡng từ cây và làm hư hại bộ rễ tạo điều kiện cho các sinh vật có hại trong đất tấn công vào bộ rễ [61]. Hầu hết các nghiên cứu hiện nay tập trung vào hai nhóm tuyến trùng ký sinh thực vật (PPN) chính là tuyến trùng bào nang (Heterodera và Globodera sp.) và tuyến trùng sần rễ (Meloidogyne sp.) gây tác động lớn đến kinh tế trên toàn thế giới.
* Đặc điểm hình thái, cấu tạo
Tuyến trùng sần rễ phân bố rộng khắp thế giới và ký sinh ở hầu hết các cây trồng quan trọng ở các vùng khí hậu khác nhau làm giảm sản lượng và chất lượng sản phẩm cây trồng. Hiện nay, có khoảng gần 80 loài ký sinh thuộc giống này, trong đó 4 loài kí sinh quan trọng nhất là: M. incognita, M. arenaria, M. javanica và M. hapla. Đây là các loài phân bố rộng và gây hại lớn ở các vùng nông nghiệp trên thế giới. Ngoài ra một số loài khác mặc dù cũng gây hại quan trọng nhưng chúng chỉ gây hại ở 1 đến 2 loại cây trồng và phân bố hẹp [62].
Vòng đời của tuyến trùng sần rễ (RKN) thay đổi từ vài ngày đến vài tháng tùy thuộc vào các yếu tố như độ ẩm, nhiệt độ, vật chủ thích hợp, loài RKN [63-64] và trải qua các giai đoạn: giai đoạn trứng, giai đoạn ấu trùng (J1, J2, J3 và J4) và giai đoạn trưởng thành (đực hoặc cái) [65].
Trong số các loài Meloidogyne, M. incognita [66] là loài phổ biến ở vùng nhiệt đới, gây hại hầu hết các loại cây trồng như dưa chuột, cà chua, thuốc lá, hồ tiêu, cà pháo, cà bát, ớt, cà phê, chuối, dứa, cây dược liệu, cây bầu bí, mướp, mồng tơi, su hào, cải bắp [63].

Hình 1.5. Chu kỳ sống của tuyến trùng sần rễ [67]
Vòng đời của tuyến trùng Meloidogyne incognita phát triển qua 5 giai đoạn chính: Trứng (ấu trùng tuổi 1) - ấu trùng tuổi 2 (ấu trùng cảm nhiễm) - ấu trùng tuổi 3 - ấu trùng tuổi 4 - tuyến trùng trưởng thành (hình 1.5) [67]. Trứng của tuyến trùng sần rễ được con cái đẻ ra ngoài trong một bọc gelatin (còn gọi là bọc trứng) nằm trên bề mặt của sần rễ. Đôi khi các bọc trứng này cũng có thể nằm bên trong nốt sần. Sau quá trình phát triển phôi thai, trứng phát triển thành ấu trùng tuổi 1 ngay bên trong trứng. Lần lột xác thứ nhất xảy ra trong trứng và phát triển thành ấu trùng tuổi 2. Trứng nở ấu trùng tuổi 2 dạng cảm nhiễm không cần sự kích thích của rễ thực vật [62]. Tuyến trùng non có dạng dài nhỏ, chiều dài thân 15 – 18 μm. Kim chích nhọn dài 10 – 12 μm, tròn như đầu kim gút. Giai đoạn ấu trùng cảm nhiễm (IJ2) là giai đoạn duy nhất lây nhiễm vào rễ cây, chúng có dạng hình giun, di chuyển và tìm rễ cây chủ bằng cách sử dụng các cơ quan cảm giác nằm trên cơ thể chúng [68].
Tuyến trùng chỉ xâm nhập vào những cây trồng thích hợp với chúng. Khi chưa gặp cây chủ thích hợp chúng có thể tồn tại một thời gian tương đối dài trong đất. Khi chuẩn bị xâm nhập vào rễ IJ2 tập trung dọc theo các tế bào non ngay tại phía sau vùng đỉnh rễ. IJ2 thường tấn công vào các mô phân sinh ở đỉnh rễ, nơi các rễ bên mọc ra tạo nên điểm xâm nhập cho các IJ2 khác và làm cho bề mặt rễ bị tổn thương. Khi IJ2
tiếp xúc với bề mặt rễ chúng thường dùng kim chích, xâm nhập ngay vào trong rễ và xảy ra ở bất kỳ phía nào của rễ. Sau khi xâm nhập vào trong rễ, tuyến trùng di chuyển giữa các tế bào vỏ rễ làm cho các tế bào bị tách dọc ra, sau đó tuyến trùng định vị tại vùng mô phân sinh của vỏ rễ và bắt đầu quá trình dinh dưỡng [62]. Khi dinh dưỡng tuyến trùng cảm phần đầu vào các tế bào mô mạch của rễ, tiết enzyme tiêu hóa làm cho quá trình sinh lý, sinh hóa của mô rễ thay đổi và hình thành các điểm dinh dưỡng cho tuyến trùng hoạt động trao đổi chất và chuyển hóa (NFS) được gọi là ‘syncytium, hoặc tế bào vận chuyển [68]. Vùng này gồm 5-6 tế bào khổng lồ (Tế bào có nhiều nhân) được tạo thành trong vùng nhu mô hoặc vùng mô libe, phía đầu tuyến trùng. Đây là sự thích nghi chuyên hóa cao của tế bào, chúng được tạo ra và duy trì bằng tuyến trùng ký sinh. Cùng với sự hình thành tế bào khổng lồ, các mô rễ xung quanh nơi tuyến trùng ký sinh cũng phình to ra tạo thành sần rễ (gall hoặc root –knot). Sần rễ thường được tạo thành trong vòng 1 ngày đến 2 ngày sau khi tuyến trùng xâm nhập. Kích thước của nốt sần liên quan đến cây chủ, số lượng IJ2 xâm nhập và loài tuyến trùng ký sinh [62].
Sau khi bắt đầu vị trí dinh dưỡng, tuyến trùng lột xác thành J3 rồi J4 và lần lột xác cuối cùng là sự biến thái thật sự đối với con đực, từ dạng cuộn gấp khúc trong IJ4 chúng được nở ra và có dạng hình giun, trong khi đó con cái có dạng hình tròn như quả lê hay quả chanh [62]. Synytium cung cấp dinh dưỡng cho sự sinh trưởng và sinh sản của tuyến trùng. Thỉnh thoảng con đực hình giun di cư ra khỏi rễ để xác định vị trí và thụ tinh cho con cái, trong khi đó, con cái phình to hình quả lê vẫn ít vận động bên trong rễ và tạo ra trứng trong chất nền gelatin. Con cái thường có màu từ trắng đến vàng, chúng sử dụng dinh dưỡng từ synytium để hỗ trợ sản xuất trứng, hầu hết trứng được đặt bên trong cơ thể con cái [64].
Tuyến trùng sần rễ có quan hệ mật thiết với các điều kiện môi trường trong đó cây chủ, nhiệt độ và các yếu tố sinh thái đất như độ ẩm, cấu trúc đất, độ thoáng khí, độ kiềm. Có thể phân biệt 2 nhóm sinh thái liên quan đến nhiệt độ là nhóm ưa nóng (Các loài điển hình như M. incognita, M. javanica, M. exigua) và nhóm ưa lạnh (Các loài điển hình như M. hapla, M. chitwoodi và có thể cả M. naasi) liên quan đến pha chuyển hóa lipid của tuyến trùng xảy ra ở 10oC. Tác hại do tuyến trùng gây ra đối với cây trồng thường có liên quan đến loại đất kiềm, là môi trường tạo ra các sốc bất lợi cho thực vật [62].