Chương 1. TỔNG QUAN
1.1 Cellulase
1.1.1. Phân loại cellulase
Cellulase là một phức hệ enzym thủy phân cellulose thông qua việc thủy phân liên kết β-1,4 glucoside hay phá vỡ liên kết hydro trong phân tử cellulose.
Cellulase có thể được chia thành 3 enzym chính dựa trên cơ chế hoạt động, trình tự axit amin và cấu trúc của chúng
Endo-(1,4)-β-D-glucanase (EC 3.2.1.4)
Exo-(1,4)-β-D-glucanase (EC 3.2.1.91)
β-glucosidase (EC 3.2.1.21).
Endoglucanase (EC 3.2.1.4) còn có tên gọi khác nhau là Endoglucanase, Endo-1,4-β-D-glucanase, Endo-1,4-glucanase, β-1,4-Endoglucan hydrolase, cellulase A, cellulosin AP, celludextrinase, carboxymethylcellulase…cơ chế hoạt động của enzym này là tấn công cellulose một cách ngẫu nhiên tại các vị trí bên trong phân tử cellulose và phân cắt chuỗi polysaccharid bằng việc chèn các phân tử nước vào giữa các liên kết β-1,4 glucosid. Kết quả tạo ra các chuỗi glucan có độ dài khác nhau. Enzym này có thể thủy phân ngẫu nhiên liên kết 1,4-β-D-glucoside giữa mạch của các cơ chất cellulose, lignin và các β-D-glucan của ngũ cốc [1].
Exoglucanase (EC 3.2.1.91) enzym này còn có tên gọi khác như Exo-β-1,4- glucanase, cellobiohydrolase, Exo-cellobiohydrolase, Exo- β-1,4-glucan cellobiohydrolase, cellobiose, C1 cellulase… bắt đầu bằng việc tấn công vào đầu khử hoặc không khử của chuỗi polysacharid và giải phóng các β-cellobiose. Exoglucanse có thể hoạt động lên các cellulose dạng tinh thể, lên lớp vỏ bên ngoài chuỗi cellulose của cấu trúc tinh thể. Đặc điểm hình học có ý nghĩa nhất của phần xúc tác của CBHs là cấu trúc hình ống được tạo ra hai lớp bề mặt. Cấu trúc hình ống này có thể toàn bộ hoặc một phần của trung tâm hoạt động. Dạng hình ống của trung tâm hoạt động có thể là hình dáng duy nhất của exoglucanase. CBHs là exoglucanse thường gặp nhất. Các CBHs khác nhau được sinh tổng hợp bởi các vi khuẩn hoặc nấm mốc với các trung tâm xúc tác phụ thuộc vào họ 5,6,7,9,48 và 74. CBHs nấm mốc thuộc họ 6, 7 và 48, CBHs vi khuẩn hiếu khí thuộc họ 6 và 48, CBHs của vi khuẩn kỵ khí thuộc họ 9 và họ 48 [1].
β-glucosidase (EC 3.2.1.21) có tên gọi khác như β-1,4-glucosidase hoạt động đặc biệt lên các cellobiose disaccharid tạo ra sản phẩm cuối glucose. Enzym này thủy phân gốc β-D-glucosid không khử ở đầu tận cùng để giái phóng glucose. β-D- glucosidase (BGs) là enzym thủy phân các cellodextrin hòa tan và các cellobiose. Hoạt tính của BGs lên các cellulose không hòa tan là không đáng kể, chúng chủ yếu
thủy phân cellobiose. Các BG khác nhau được thu nhận từ các vi khuẩn, nấm mốc, động vật và thực vật khác nhau với trung tâm xúc tác phụ thuộc vào họ 1,3 hoặc 9. Dựa trên các dữ liệu phân tích cấu trúc tương đồng, cấu trúc lập thể hóa học của họ 1 và 3 BGs thuộc loại duy trì, trong khi họ 9 thuộc loại đảo ngược. BGs có trung tâm hoạt động dạng túi, nơi chúng được liên kết với các đầu gluocse không khử và ghim các glucose từ cellobiose và cellodextrin [1].
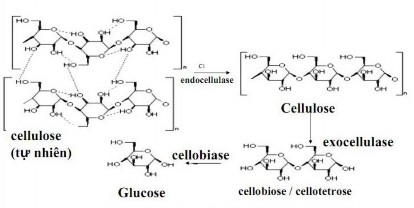
Hình 1.1 Quá trình phân giải cellulose của cellulase [2]
Trong hầu hết các cách phân loại cơ bản thì thì cellulase thuộc nhóm enzym đơn chức năng được xác định đó là các cellobiohydrolase (CBHs, exo-β-(1,4)-d glucanases), endoglucanase (EGs, endo β-(1,4)-glucanase) và β-d-glucosidase. CBHs là enzym thủy phân cellulose từ đầu chuỗi, trong khi EGs là loại enzym thủy phân cellulose một cách ngẫu nhiên từ bên trong chuỗi. Cellobiase thủy phân các cellobiose, sản phẩm cuối cùng của CBHs là glucose do vậy mà ngăn cản được sản phẩm cuối kìm hãm enzym [3].
EGs có khả năng tạo ra các cellulose chuỗi ngắn hơn để thuận tiện cho quá trình thủy phân bởi enzym. Sự kết hợp hoạt tính của EG và CBHs giải phóng một lượng lớn các cellodextrin hòa tan và các cellobiose. β-d-glucosidase thủy phân các dimer glucose thành glucose
.
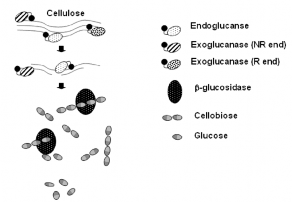
Hình 1.2 Cơ chế thủy phân cellulose của hệ cellulose [4]
1.1.2. Nguồn thu nhận cellulase
Cellulase được sinh tổng hợp trong tự nhiên bởi một số loài vi sinh vật và cả một số động vật và côn trùng. Các vi sinh vật sinh cellulase có mặt ở nhiều nơi, những vi sinh vật này đóng vai trò quan trọng trong việc chuyển hóa các polysacharid phức tạp thành các đường đơn giản mà chúng có thể đồng hóa được. Các vi sinh vật có khả năng sinh cellulase trong tự nhiên bao gồm cả xạ khuẩn, nấm mốc và vi khuẩn. Các vi khuẩn có khả năng phân giải cellulose có cả vi khuẩn hiếu khí, kỵ khí, cả chủng ưa ấm và chịu nhiệt. Môi trường mà chúng có mặt rất đa dạng bao gồm cả những nơi có nhiệt độ, áp suất cao tới những nơi có pH khắc nghiệt [5].
Vi khuẩn sinh cellulase đã từng được phân lập từ rất nhiều nguồn khác nhau như dạ cỏ ở các động vật nhai lại (Hungater, 1957), đất (Black et al., 1985) phân (Madden, 1982), chất thải đô thị (Stutzenberger et al., 1970), nước cống (Muray et al., 1984), suối nước nóng (Sissons et al., 1987) [6]
Côn trùng là động vật được quan tâm nhiều nhất về nhóm vi sinh vật cộng sinh trong đường ruột và sự đa dạng sinh học của chúng. Có khoảng hơn 20% côn trùng có liên quan tới vi sinh vật cộng sinh. Vai trò của các vi sinh vật cộng sinh này là khả năng sinh cellulase giúp cho các côn trùng có thể chuyển hóa các nguồn thức ăn có nguồn gốc là lignocellulose (Morrison et al,2009) [7].
Từ những năm 1992 Hethener và cộng sự đã nghiên cứu phân lập vi khuẩn sinh cellulase từ ruột của mối ăn gỗ Nasutitermes lujae. Kết quả đã phân lập được chủng vi khuẩn yếm khí Clostridium termitidis sp đồng thời nghiên cứu quá trình sinh trưởng và phát triển của chủng.
Đến nay đã có rất nhiều vi khuẩn từ ruột mối được chứng minh có khả năng sinh cellulase như chủng MX5T được phân lập từ ruột mối Mastotermes ở Autrialia được xác định là có khả năng sinh cellulase và xylanase, với kỹ thuật định tên bằng 16S rDNA chủng MX5T được xác định là Cellulosimicrobium variable [8].
Các chủng vi khuẩn hiếu và yếm khí có khả năng sinh cellulase cũng đã được phân lập từ mối Zootermopsis angusticollis được công bố bới Wenzel và cộng sự 2001 là các chủng thuộc chi Cellulomonas, Bacillus, Pseudomonas, Rhizobium [9].
Các vi khuẩn phân lập từ loài mối Heterotermes và Odontotermes được xác định là có tiềm năng sinh cellulase, kết quả định tên cho thấy các chủng phân lập được thuộc các chi Bacillus, Staphylococus và Enterobacter. Bacillus cereus được xác định là có tiềm năng sinh cellulase cao nhất với hoạt độ endoglucanase, FPase và β-glucosidase tương ứng là 5,06 U/mg; 2,52 U/mg và 6,01 U/mg sau khi đã tối ưu hóa môi trường nuôi cấy theo Chen và cộng sự [10].
1.1.3. Sinh tổng hợp cellulase từ vi khuẩn
Sinh tổng hợp cellulase từ vi khuẩn phụ thuộc rất nhiều vào các thông số của môi trường nuôi cấy như nhiệt độ, nguồn dinh dưỡng, tỷ lệ oxy, thời gian nuôi cấy, thành phần môi trường, pH của môi trường …
1.1.3.1 Sinh tổng hợp cellulase từ vi khuẩn
Ảnh hưởng nhiệt độ
Nhiệt độ ảnh hưởng đến sự phát triển và khả năng sinh tổng hợp enzym của vi sinh vật cũng như tính chất của enzym được tổng hợp. Sinh trưởng và sinh tổng hợp enzym thường bị kìm hãm nhanh chóng ở nhiệt độ cao hơn nhiệt độ tối ưu. Nhiệt độ quá cao gây ra sự biến tính enzym, các protein vận chuyển và các protein khác, màng tế bào bị tổn thương vì hai lớp lipid bị hòa tan [11]. Tuy nhiên, điều kiện nhiệt độ rất thấp, màng tế bào có thể bị kết đông, enzym ngừng hoạt động. Nói chung, nếu vượt quá nhiệt độ tối ưu của vi sinh vật, chức năng và kết cấu tế bào đều bị ảnh hưởng. Do ảnh hưởng hai mặt, vừa có lợi vừa có hại của nhiệt độ đối với vi sinh vật mà có thể xác định các khoảng nhiệt độ cơ bản đối với sự sinh trưởng của vi sinh vât là nhiệt độ thấp nhất (minimum), nhiệt độ tối ưu (optimum) và nhiệt độ cao nhất (maximum) đối với sự sinh trưởng [12].
Nhiệt độ tối ưu cho sinh trưởng Micrococus sp là 37oC nhưng nhiệt độ tối ưu
cho quá trình sinh cellulase là 27oC. Nhiệt độ tối ưu cho quá trình sinh cellulase là 30oC đối với các chủng Cellulomonas ASN2 đối với các chủng phân lập từ đất và khả năng sinh tổng giảm khi nhiệt độ lên 45oC (M. Irfan et al.,2012)[13]
Với vi khuẩn phân lập từ ruột mối đã được báo cáo có nhiệt độ tối ưu cho quá trình sinh cellulase là 37oC (D.Submitted et al, 2013) [14] và (Apurv et al., 2014) [15].
Ảnh hưởng của pH
Phần lớn vi sinh vật phát triển tốt nhất trong môi trường thuộc vào dải pH trung tính 6,5 – 7,5. Nhưng trong quá trình sinh trưởng của vi khuẩn, một số axit hữu cơ được giải phóng vào môi trường, làm giảm pH của môi trường và do đó gây trở ngại hoặc ức chế hoàn toàn sự phát triển của vi sinh vật. Ion H+ nằm trong thành phần môi trường tùy theo nồng độ của chúng mà làm tăng hoặc giảm khả năng thẩm thấu của tế bào đối với những ion nhất định. Ngoài ra, chúng cũng có thể làm ức chế phần nào các enzym có mặt trên thành tế bào. Vi sinh vật có thể chỉ tồn tại hoặc
thậm chí chết trong môi trường axit hay kiềm. Đối với vi khuẩn, thuận lợi nhất là chúng phát triển trong môi trường trung tính, axit yếu hoặc kiềm yếu [16].
pH trong ruột mối bậc thấp ở khoảng trung tính 6,0-7,6 còn ở mối bậc cao thì có pH cao hơn (8-9), thay đổi tùy vào vị trí ruột giữa, ruột trước, ruột sau. Các loài mối bậc cao được báo cáo có pH trong ruột sau lên đến 12 (Cubitermes severus và Procubitermes aburiensis) [17].
pH là một trong những yếu tố quan trọng cho quá trình sinh tổng hợp cellulase. Giá trị pH axit hay bazơ quá cao đều ảnh hưởng tới quá trình sinh tổng hợp cellulase. pH thích hợp nhất cho quá trình sinh enzym là giá trị pH môi trường mà tại đó enzym thu nhận được có hoạt tính cao nhất [14].
Ảnh hưởng của thời gian nuôi cấy
Thời gian lên men có ảnh hưởng khá mạnh tới sự sinh trưởng và sinh tổng hợp enzym của chủng. Kéo dài thời gian lên men có thể làm giảm hoạt độ của enzym do suy giảm các chất dinh dưỡng và tạo ra các sản phẩm phụ trong môi trường lên men. Thời gian nuôi cấy thích hợp của mỗi loại vi sinh vật được xác định bằng thời gian cho phép tích tụ lượng enzym tối đa, thời gian đó phụ thuộc chủ yếu vào chủng vi sinh vật và thành phần môi trường nuôi cấy. Quá trình sinh trưởng của vi sinh vật trong hệ kín trải qua bốn pha liên tiếp là pha tiềm phát (pha lag), pha lũy thừa, pha cân bằng, pha suy vong [7].
Trong một nghiên cứu thu nhận và xác định tính chất của cellulase và xylanase từ C.cellulans, các tác giả nhận thấy cellulase được tạo ra nhiều nhất sau 24 giờ nuôi cấy và duy trì lượng tương đối ổn định trong khoảng thời gian khoảng 70 giờ tiếp theo [18]. Tuy nhiên thời gian sinh tổng hợp cũng phụ thuộc nhiều vào thành phần môi trường và bản chất của cơ chất sử dụng [18].
Quá trình sinh tổng hợp cellulase từ nấm mốc thường kéo dài hơn so với từ vi khuẩn. Theo Gupta et al (2015) các chủng Asperillus sinh cellulase tốt nhất sau 72 giờ nuôi cấy. Với Trichoderma thì quá trình sinh tổng hợp tốt nhất sau 5 ngày nuôi cấy. Với cellulase thu nhận từ vi khuẩn có hoạt tính cao nhất sau 24-72 giờ
nuôi cấy. Thời gian thu nhận cellulase từ vi khuẩn thường 48- 72 giờ đối với các chủng vi khuẩn thuộc chi Bacillus, Pseudomonas, Cellulomonas… .
Ảnh hưởng của tốc độ lắc
Chế độ lắc trong lên men là một trong những yếu tố thúc đẩy nhanh quá trình lên men. Tốc độ lắc phù hợp giúp rút ngắn thời gian và làm tăng hiệu quả quá trình lên men đối với các vi sinh vật hiếu khí. Lượng oxy ảnh hưởng tới quá trình trao đổi chất và tích lũy các sản phẩm sinh tổng hợp của vi sinh vật [19].
Theo công bố của Liu và cộng sự, khi nuôi cấy Cellulosimicrobium thì việc sục khí và khuấy trộn là hết sức cần thiết. Oxy là yếu tố cơ bản ảnh hưởng đến sự sinh trưởng phát triển và quá trình trao đổi chất của tế bào. Trong một số nghiên cứu với các chủng thuộc chi Cellulosimicrobium, tốc độ lắc sử dụng nằm trong khoảng từ 150 vòng/phút - 180 vòng/phút [12].
1.1.3.2 Ảnh hưởng của nguồn dinh dưỡng
Nguồn dinh dưỡng cacbon
Hầu hết các nghiên cứu nuôi cấy vi khuẩn sinh cellulase đều cung cấp nguồn cacbon trong môi trường bằng cách sử dụng các loại đường khác nhau. Các nguồn đường được sử dụng rất đa dạng tùy thuộc vào từng chủng và mục đích nuôi cấy như glucose, galactose, sucrose, maltose, mannose, tinh bột, cellulose và các chất có chứa nguyên tử C. Trong một so sánh về các loài trong chi Cellulosimicrobium, các tác giả nhận thấy loại đường chính trong cấu tạo thành tế bào vi khuẩn là galactose [8]. Thay đổi các điều kiện nuôi cấy như thêm 0,5% các nguồn cacbon (tinh bột, maltose, glucose, surose, bột cellulose cho thấy Cellulomonas có khả năng sử dụng nhiều nguồn cacbon khác nhau, với nguồn cacbon glucose làm tăng hoạt tính cellulase đạt 0,475±0,019IU/ml/min [16].
Nghiên cứu của Jaradat và cộng sự cho thấy khi sử dụng các nguồn cacbon khác nhau trong môi trường nuôi cấy, cũng đã xác định glucose là nguồn cacbon thích hợp nhất cho quá trình sinh tổng hợp cellulase của chủng Streptomyces sp [20].
Nghiên cứu ảnh hưởng nguồn dinh dưỡng cacbon của vi khuẩn sinh cellulase từ ruột mối phân lập được bởi Bholay và cộng sự cho thấy glucose, fructose, xylose và tinh bột làm tăng khả năng sinh cellulase với các chủng Pseudomonas phân lập được từ ruột mối [15].
Nguồn dinh dưỡng nitơ
Nguồn Nitơ là nguồn dinh dưỡng có chứa nitơ là thành phần xây dựng protein và protoplasm. Với các nguồn nitơ sử dụng trong nghiên cứu cho quá trình sinh tổng hợp cellulase thường là các amino axit như alanine, glutamic acid, cystine,
lysine, tyrosine, các nguồn nitơ vô cơ (NH4)2SO4, KNO3, amonium citratat, NH4Cl, NaNO3 hay các nguồn nitơ hữu cơ như peptone, trytone, yeast extract và ure.
Nghiên cứu trên các chủng ASN2 cho thấy NH4Cl và pepton cho hoạt tính cellulase cao nhất là 0,45 và 0,47 U/ml/min ở 35oC trong 48h. (báo cáo của Jaradat et al) [20]
Theo nghiên cứu của Vyas et al thì (NH4)2SO4 và pepton là nguồn nitơ thích hợp nhất cho với quá trình sinh endocellulase và exocellulase khi nuôi cấy chủng vi khuẩn thuộc chi B.subtilis trên môi trường lỏng [21].
Các chủng Pseudomonas sp phân lập từ ruột mối bởi Bholay và cộng sự thì Amoni nitrat và Kali nitrat làm tăng khả năng sinh tổng hợp cellulase (Bholay et al, 2014) [15]. Amino acid cũng ảnh hưởng tới quá trình sinh tổng hợp cellulase và theo báo cáo của Sharma et al thì alanine, L-asparagin cũng cho thấy làm tăng quá trình sinh tổng hợp cellulase với chi thuộc Bacillus và Pseudomonas [22].
Nguồn cơ chất
Trong quá trình sinh tổng hợp cellulase thì việc sử dụng CMC, giấy lọc và một số nguồn cellulose khác như là nguồn dinh dưỡng đồng thời là chất cảm ứng quá trình sinh tổng hợp cellulase. Nồng độ cơ chất cellulose cũng đóng vai trò quan trọng cho quá trình sinh tổng hợp cellulase. Các nghiên cứu thường sử dụng các nồng độ từ 0,5% - 5% cơ chất để khảo sát nồng độ tối ưu cho quá trình sinh tổng hợp. (Nisha, 2015) [16]. Theo báo cáo của Das et al, lại cho thấy sử dụng CMC là cơ chất cho quá trình sinh tổng hợp nhưng cũng nguồn cacbon duy nhất lại là tốt nhất cho qúa trình sinh tổng hợp cellulase với chủng Bacillus sp (3,028µm/mg/min) [23].
Các chất thải nông nghiệp chi phí thấp như bã mía, lõi ngô, vỏ khoai tây, vỏ đậu,… được xác định là nguồn cơ chất tốt đối với sinh trưởng và sinh tổng hợp từ
C. cellulans [12]. Trong nghiên cứu của Jian-Min Song và cộng sự, bã mía được sử dụng như nguồn cacbon chính trong môi trường khoáng cơ bản M9 cũng cho hoạt độ cellulase và xylanase tương đối cao [18].
Ngoài vai trò là nguồn dinh dưỡng đối với vi sinh vật, việc sử dụng các cơ chất rắn còn trực tiếp thể hiện tiềm năng phân hủy sinh khối lignocellulose, góp phần tận dụng hiệu quả nguồn thải từ các ngành nông nghiệp [24].
1.1.4. Đặc tính của enzym cellulase từ vi khuẩn
Các đặc tính của cellulase rất đa dạng tùy thuộc vào nguồn thu nhận. Enzym thu nhận từ các nguồn khác nhau thì các thông số như pH tối ưu, nhiệt độ tối ưu, khả năng chịu pH, nhiệt độ hay ảnh hưởng của các yếu tố hóa học, vật lý lên hoạt độ enzym cũng khác nhau [25].
1.1.4.1. pH tối ưu và nhiệt độ tối ưu của enzym
Hầu hết cellulase được nghiên cứu có sự giống nhau về pH tối ưu, độ hòa tan và thành phần amino acid nhưng độ bền nhiệt và độ đặc hiệu cơ chất có thể thay đổi. Tuy nhiên, một điều đáng lưu ý là các enzym nghiên cứu còn có thể có các hoạt tính khác ngoài hoạt tính cellulase và đặc điểm này có thể ảnh hưởng đến chế phẩm sản xuất. Các chế phẩm cellulase hoạt động hiệu quả trong khoảng pH 3-7, pH tối ưu thường nằm trong khoảng 5-6, nhiệt độ tối ưu từ 40-50°C [26].
Hoạt độ của cellulase tạo thành từ vi sinh vật chịu sự ảnh hưởng của rất nhiều yếu tố. Các nghiên cứu được tiến hành đã xác định ảnh hưởng của nồng độ cơ chất, thời gian phản ứng enzym, tác động của các ion kim loại và chất hóa học,… đến hoạt độ enzym. Tuy nhiên, pH, nhiệt độ tối ưu và độ bền nhiệt, bền pH của enzym là một trong các đặc tính quan trọng có tính ứng dụng cao nên đa số các nghiên cứu tập trung xác định các yếu tố này [27].
pH và nhiệt độ có ảnh hưởng lớn đến hoạt độ và độ bền của enzym cellulase. Trong nghiên cứu về tính chất của cellulase và xylanase sinh tổng hợp bởi Cellulosimicrobium cellulans trên môi trường M9 có bổ sung bã sắn của Jian-Min Song và Dong-Zhi Wei, cellulase tạo thành có pH tối ưu là 5, giữ được hoạt tính sau 8h ở nhiệt độ 30°C và 64% hoạt tính sau 1h ở 45°C. Các enzym bền nhiệt được chia thành 3 nhóm tùy thuộc vào khoảng nhiệt độ bền của enzym: enzym có độ chịu nhiệt vừa phải (moderately thermostable) (45-65°C), enzym bền nhiệt (thermostable) (65-85°C) và enzym bền nhiệt cao (extremely thermostable) (>85°C). Do đó, cellulase và xylanase tạo bởi C.cellulans có thể được phân loại thuộc nhóm chịu nhiệt vừa phải. Về pH, nghiên cứu chỉ ra, cellulase tạo thành giữ được hoạt tính tốt nhất ở pH 5 và hoạt tính suy giảm mạnh ở pH axit <3,4 [18].
Bảng 1.1: Đặc tính của cellulose và Xylanase từ C.cellulans [18]
Cellulase | Xylanase | |
pH tối ưu | 5,0 | 5,6 |
Nhiệt độ tối ưu (ºC) | 50 | 50 |
Hoạt độ (IU/ml) | 0,03 ±0,001 | 0,7±0,02 |
Có thể bạn quan tâm!
-

 Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 1
Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 1 -
 Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 2
Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 2 -
![Tính Bền Nhiệt Của Cellulase Và Xylanase Từ C.cellulans [18].](https://tailieuthamkhao.com/uploads/2023/02/19/nghien-cuu-cellulase-tu-vi-khuan-ruot-moi-phan-lap-o-viet-nam-4-1-120x90.jpg) Tính Bền Nhiệt Của Cellulase Và Xylanase Từ C.cellulans [18].
Tính Bền Nhiệt Của Cellulase Và Xylanase Từ C.cellulans [18]. -
 Ứng Dụng Giải Trình Tự Genome Để Phát Hiện Tiềm Năng Vi Khuẩn Sinh Tổng Hợp Cellulase Trong Phân Giải Lignocellulose
Ứng Dụng Giải Trình Tự Genome Để Phát Hiện Tiềm Năng Vi Khuẩn Sinh Tổng Hợp Cellulase Trong Phân Giải Lignocellulose -
 Phương Pháp Phân Lập Và Tuyển Chọn Chủng Sinh Tổng Hợp Cellulase
Phương Pháp Phân Lập Và Tuyển Chọn Chủng Sinh Tổng Hợp Cellulase