điều đó dẫn tới hiệu suất thủy phân thấp khi sử dụng enzym từ G4 để thủy phân rơm và bã mía (Hình 3.14). Hơn nữa khi quan sát quá trình thủy phân giấy lọc có thể thấy enzym từ MP1 thủy phân giấy lọc tốt hơn enzym từ G4 (Hình 3.21).

Hình 3.21. Thử nghiệm phân giải giấy lọc của MP1 và G4
Chế phẩm cellulase thu được từ Cellulosimicrobium sp. MP1 có khoảng pH hoạt động rộng hơn và có khả năng hoạt động tốt ở pH kiềm, rất phù hợp để thủy phân nguyên liệu cellulose.
Thử nghiệm với các cơ chất cho thấy chủng MP1 thuộc chi Cellulosimicrobium phân lập được có khả năng phân hủy các cơ chất bã mía, rơm, giấy lọc (hình 3.21), cho thấy chủng MP1 phân lập được có khả năng tiết ra hỗn hợp các enzym phá hủy được cấu trúc phức tạp của cơ chất. Hỗn hợp này (enzymatic bulk) có thể bao gồm endoglucanases, cellobiohydrolases và β-glucosidases để thủy phân thành tế bào của nguyên liệu, cũng như có chứa endoxylanases và β- xylosidases đóng vai trò thủy phân các thành phần hemicellulose.
Vi những lý do trên chủng Cellulosimicrobium sp. MP1 sẽ được tiếp tục nghiên cứu ở mức độ AND.
3.5. Xác định đặc tính di truyền và các gen mã hóa enzym thủy phân cellulose của chủng C. cellulans MP1
3.5.1. Trình tự gen và một số đặc điểm khái quát của chủng C. cellulans MP1
Vì phân tích trình tự gen đã và đang giúp mở ra con đường tìm hiểu nhiều hơn về các enzym liên quan đến sự phân giải sinh khối thực vật, trình tự gen của chủng C. cellulans MP1 đã được phân tích với kỹ thuật Illumina Miseq. Thư viện gen thu được đã cho ra tất cả là 443 986 169 nucleotit và được tổng hợp để tạo thành 4 487 842 bản đọc với vị trí cụ thể và với độ lặp là 92,2 lần. Trình tự gen của chủng MP1 (4 580 223 bp với hàm lượng GC 73.9%) được sắp xếp lại thành một nhiễm sắc thể hình tròn với 23 contig và không phát hiện thấy có plasmid (hình 3.22). Toàn bộ hệ gen của chủng được dự đoán có 4117 gen bao gồm 4 046 trình tự
mã hóa protein (CDS), 67 trình tự quy định rRNA, 3 trình tự quy định tRNA, 1 trình tự quy định tmRNA (hình 3.22).
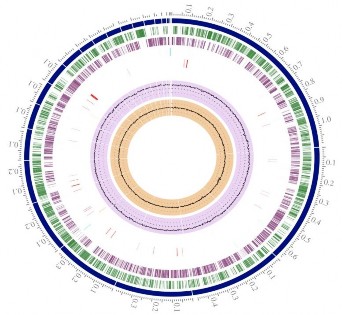
Hình 3.22 Sơ đồ genom của C.cellulans MP1
3.5.2. Các cụm gen và chức năng gen của chủng MP1
Trong toàn bộ các trình tự mã hóa protein thì 79.5% (3216 trong 4046) được phân vào 21 trong 25 nhóm chức năng COG. Trong đó, các nhóm chiếm nhiều nhất là: phiên mã (K: 365 trình tự), vận chuyển và chuyển hóa carbonhydrate (G: 307), vận chuyển và chuyển hóa amino acid (E: 224), sản sinh và chuyển hóa năng lượng (C: 199), và vận chuyển và chuyển hóa (P: 180). Ngược lại, nhóm liên quan đến việc điều hòa chu trình, phân chia tế bào và nhiễm sắc thể (D: 33), cấu tạo và điều khiên sợi nhiễm sắc (B: 2) và di chuyển tế bào (N: 1) là các nhóm chiếm ít nhất.
Việc phân tích chức năng của các gen cung cấp cái nhìn sâu hơn về các chức năng của chủng MP1. Tổng số 3165 gen được phân vào 43 lớp con bao gồm 13 lớp con thuộc lớp cấu tạo tế bào (CC), 10 lớp con thuộc lớp liên quan đến phân tử chức năng (MF), và 20 lớp con liên quan đến các quá trình sinh học (BP). Cụ thể, lớp CC chiếm nhiều gen nhất (1638 gen, 45,3%) tiếp theo là lớp BP (1424 gen, 39,4%) và MF ( 553 gen, 15,3%). Các con đường chiếm nhiều nhất là tế bào (GO:0005623; 508 gen), thành phần tế bào (GO:0044464; 379 gen) và cấu tạo thành tế bào (GO:0016020; 340gen). Trong lớp BP 3 nhóm chiếm ưu thế là quá trình chuyển hóa (GO:0008152; 378 gen), quá trình trong tế bào (GO:0009987; 379) gen và phát triển (GO:0040007; 281 gen). Trong các lớp nhỏ thuộc vào lớp MF, các gen thuộc nhóm có hoạt tính xúc tác (GO:0003824) và bám (GO:0005488) chiếm 311 và 176 gen.
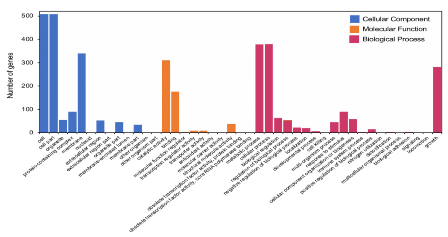
Hình 3.23 Phân loại chức năng gen
3.5.3. So sánh COG toàn bộ hệ gen và các enzym liên quan đến chuyển hóa carbonhydrate
Sự tiến hóa của vi khuẩn là nhờ sự tiến hóa thẳng và bắt nguồn từ 1 gen tổ tiên duy nhất, từ đó tạo thành các gen tiến hóa thẳng trong các loài khác nhau, để có thể hiệu rõ hơn về chức năng, cấu tạo cũng như sự biến đổi của các gen thì cần phải có sự so sánh các nhóm COG ở các chủng khác nhau. Các nhóm COG của chủng
C. cellulans MP1 được so sánh với của 4 chủng khác gồm J36, LMG16121, và JKA48. Chủng C. cellulans MP1 có 3405 gen thuộc vào các nhóm COG và 794 gen độc lập. Các chủng J36 và ZKA48 có lần lượt 3604 và 3605 gen thuộc các nhóm COG, trong khi chủng LMG16121 có ít gen COG nhất (hình 3.25). Trong đó thì số gen đơn lẻ lớn nhất thuộc về chủng MP1. Biểu đồ Venn Edward 6 chiều cho thấy rằng có 2539 gen COG được tìm thấy trong cả 4 chủng thuộc chi C. cellulans. Trong số các COG riêng biệt của mỗi chủng thì chủng MP1 có 20 và là số lượng lớn nhất trong 4 chủng. Để kiểm chứng kết quả này, công cụ xây dựng cây phân loài REALPHY đã được sử dụng. Với lựa chọn cây có khả năng xảy ra cao nhất (maximum likelihood), cây phân loài được xây dựng cho thấy 2 chủng MP1 và LMG16121 tạo thành một nhánh riêng, cho thấy một mối quan hệ tiến hóa mật thiết (hình 3.24).
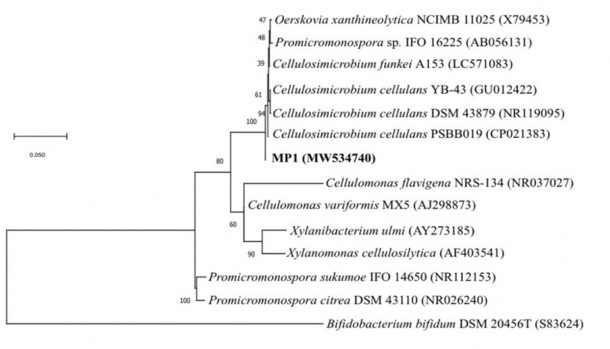
Hình 3.24. Cây phân loài chủng C.cellulans MP1
Sử dụng trang web dbCAN2, các gen được dự đoán tổng hợp các CAZy của chủng C. cellulans MP1 được sàng lọc để tìm ra các gen liên quan đến việc phân hủy cellulose và hemicellulose. Sau khi loại bỏ các trình tự không liên quan, 195 vùng gen được dự đoán là CAZy được tìm thấy, chúng chiếm 4,8% trong tổng số 3216 vùng của chủng này. Các enzym glycoside hydrolase (GH) phụ trách việc phân hủy phần lớn sinh khối thực vật (cellulose và hemicellulose) được dự đoán là họ enzym nhiều nhất và chiếm đến 99 enzym. Tiếp theo là 39 enzym glycosyl transferase (GT), 37 đơn vị bám vào carbohydrate (CBM), 11 enzym carbohydrate esterase (CE), 6 enzym với các hoạt tính phụ trợ (AA) và 3 enzym polysaccharide lyases (PL) được phát hiện
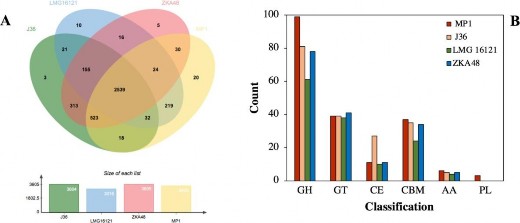
Hình 3.25 So sánh genom của MP1 với các chủng C.cellulans
Hệ gen của 4 chủng C. cellulans MP1, J36, LMG16121, ZKA48 đã được so sánh. Về cơ bản, số lượng vùng thuộc nhóm CAZy của chủng MP1 lớn hơn rất nhiều so với các chủng khác (hình 3.25). Các vùng CAZy được tìm thấy trong C. cellulans MP1 có độ tương đồng cao với chủng C. cellulans J36. Mặc dù 2 chủng MP1 và LMG16121 có quan hệ gần gũi, nhưng MP1 sở hữu nhiều hơn chủng LMG16121 56 vùng CAZy. Ngoài chủng LMG16121 các số vùng thuộc vào các họ GT, CBM và AA của các chủng còn lại gần như là bằng nhau, cho thấy sự tác động đồng thời của các gen vùng này trong quá trình phân hủy sinh khối thực vật
3.5.4. Khai thác các enzym thủy phân sinh khối thực vật từ nguồn gen của chủng C. cellulans MP1.
Phân tích các gen CAZy đã tìm ra 30 gen liên quan đến cellulose trong hệ gen của C. cellulans MP1. Các họ enzym lớn, liên quan đến việc thủy phân cellulose bao gồm GH6, GH9, GH48, GH10, GH16, GH1, GH3, GH13, và GH64.
Việc định danh gen đã phát hiện 5 endoglucanase (2 thuộc họ GH6 và 3 thuộc họ GH9), 3 exoglucanases (GH6,GH10, GH48) và lichenase (GH16) tồn tại trong hệ gen của chủng MP1 (bảng 3.17). Trong 8 gen tổng hợp endo và exo glucanase thì 5 gen (Orf_454, Orf_1616, Orf_3244, Orf_1607, Orf_1611) có gắn thêm vùng CBM2 (một vùng có thể bám vào phần tinh thể của các cơ chất cellulose, chitin, xylan). CBM2 là vùng CBM có mặt nhiều nhất trong hệ gen của MP1. Hệ gen của MP1 còn có các gen quy định 3 enzym thuộc GH1 và GH6, enzym thuộc GH3 có hoạt tính β- glucosidase được xem là góp phần quan trọng trong quá trình thủy phân cellulose và 3 enzym khác thuộc họ GH13 có hoạt tính α-glucosidase. Ngoài ra còn có 2 họ enzym polysaccharide lyase liên quan đến việc phân giải pectin: PL4 (Orf_102) và PL1 (Orf_1326) (Bảng 3.17).
Bảng 3.17 Danh mục các cellulase và hemicellulase dự đoánbiểu hiện trên hệ gen của
C. cellulans MP1
Locus tag | Dự đoán chức năng | |
Cellulose- related | Orf_454, Orf_1616, Orf_2130, Orf_2755, Orf_3244 | Endoglucanase [EC 3.2.1.4] |
Orf_1607, Orf_1610, Orf_1611 | Exoglucanase [EC 3.2.1.91] | |
Orf_2289 | Lichenase [EC 3.2.1.73] | |
Orf_2294, Orf_2388, Orf_2464, Orf_2606, Orf_3385, Orf_401, Orf_403, Orf_404, Orf_922 | β-glucosidase [EC 3.2.1.21] | |
Orf_2704, Orf_2893 | Oligo-1,6-glucosidase [EC 3.2.1.10] | |
Orf_2898 | Maltodextrin glucosidase [EC 3.2.1.20] | |
Orf_802 | Glucan endo-1,3-β-glucosidase [EC 3.2.1.39] | |
Orf_3703 | β-galactosidase [EC 3.2.1.23] | |
Orf_2449 | Trehalose-6-phosphate hydrolase [EC 3.2.1.93] | |
Orf_102 | Pectate trisaccharide-lyase [EC 4.2.2.22] | |
Orf_1326 | Pectate lyase [EC 4.2.2.2] | |
Orf_19, Orf_21 | Levanase [EC 3.2.1.80] | |
Orf_20 | Levanbiose-producing levanase [EC 3.2.1.64] | |
Orf_22 | Levansucrase [EC 2.4.1.10] | |
Hemicellulose- related | Orf_905 | Mannan endo-1,4-β-mannosidase [EC 3.2.1.78] |
Orf_3988, Orf_4004 | Bifunctional β-xylosidase/α- arabinosidase [EC 3.2.1.37; EC 3.2.1.55] | |
Orf_4003 | Arabinoxylan arabinofuranohydrolase [EC 3.2.1.55] | |
Orf_35, Orf_2605 | α-xylosidase [EC 3.2.1.177] | |
Orf_24, Orf_2083, Orf_3386, Orf_3999, Orf_4000, Orf_4002 | α-L-arabinofuranosidase [EC 3.2.1.55] | |
Orf_34, Orf_3650, Orf_4001 | Non-reducing end β-L- arabinofuranosidase [EC 3.1.1.185] | |
Orf_3698 | Exo-α-(1->6)-L-arabinofuranosidase [EC 3.2.1.-] | |
Orf_1000, Orf_2117, Orf_3772 | Endo-1,4- β-xylanase [EC 3.2.1.8] | |
Orf_266, Orf_2247 | α-galactosidase [EC 3.2.1.22] |
Có thể bạn quan tâm!
-

 Đặc Điểm Hình Thái, Sinh Lý Của 11 Chủng Vi Khuẩn Tuyển Chọn
Đặc Điểm Hình Thái, Sinh Lý Của 11 Chủng Vi Khuẩn Tuyển Chọn -
 Bề Mặt Đáp Ứng Của Quá Trình Sinh Tổng Hợp Cmcase Từ Bacillus G4 Cho Thấy Sự Tương Tác Giữa (A) Hàm Lượng Bột Đậu Tương Và Tinh Bột . (B) Hàm
Bề Mặt Đáp Ứng Của Quá Trình Sinh Tổng Hợp Cmcase Từ Bacillus G4 Cho Thấy Sự Tương Tác Giữa (A) Hàm Lượng Bột Đậu Tương Và Tinh Bột . (B) Hàm -
 Ảnh Hưởng Của Môi Trường Nuôi Cấy Tới Khả Năng Sinh Enzym
Ảnh Hưởng Của Môi Trường Nuôi Cấy Tới Khả Năng Sinh Enzym -
 Ảnh Hưởng Của Tiền Xử Lý Cơ Chất Tới Hiệu Suất Đường Hóa
Ảnh Hưởng Của Tiền Xử Lý Cơ Chất Tới Hiệu Suất Đường Hóa -
 Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 14
Nghiên cứu cellulase từ vi khuẩn ruột mối phân lập ở Việt Nam - 14 -
 Đặc Điểm Các Chủng Vi Khuẩn Phân Lập Được Trên Đĩa Thạch Từ Các Mẫu Mối 01- 04
Đặc Điểm Các Chủng Vi Khuẩn Phân Lập Được Trên Đĩa Thạch Từ Các Mẫu Mối 01- 04
Xem toàn bộ 138 trang tài liệu này.
Xét về khả năng thủy phân hemicellulose, một số protein được xác định là có tham gia vào quá trình này. Các họ enzym lớn nhất tham gia vào quá trình phá vỡ cấu trúc của hemicellulose là GH43, GH31, GH127, GH51, GH10, GH36, và GH4, và tổng số các enzym được tìm thấy là 21. Trong số đó, chiếm nhiều nhất là GH43 (9 enzym), GH124 (3 enzym) và GH31 (2 enzym), điều này cho thấy chủng MP1 có thể có một tiềm năng lớn trong việc thủy phân và cắt các mạch nhánh trong cấu trúc hemicellulose. Các gen này được dự đoán bao gồm: mannan endo-1,4-β-
mannosidase, arabinoxylan arabinofuranohydrolase, α-xylosidase, α-L- arabinofuranosidase, β-L-arabinofuranosidase, endo-1,4-β-xylanase, α- galactosidase, và enzym đa chức năng β-xylosidase/α-arabinosidase. Chỉ có 2 enzym là mannan endo-1,4-β-mannosidase và α-galactosidase đã được chứng minh về khả năng thủy phân mannan, thành phần quan trọng thứ 2 trong hemicellulose. 2 enzym endo-1,4-β-xylanase (Orf_100 và Orf_3772) và 1 enzym đa chức năng β- xylosidase/α-arabinosidase (Orf_3988) được gắn thêm CBM2 và CBM9. Một điều hiếm thấy là các gen mã hóa các enzym arabinofuranosidase (orf_3999, orf_4000, orf_4001 và orf_4002), arabinoxylan arabinofuranohydrolase (orf_4003), và enzyme đa chức năng β-xylosidase/α-arabinosidase (orf_4004) lại được nhóm với nhau và nằm trong cùng 1 operon.
Tương tự trong nghiên cứu của Chen và cộng sự (2019), ở nghiên cứu này vi khuẩn Micromonospora sp CP22 cũng có khả năng sinh cellulase, xylanase và mananase được phân lập từ phân ủ của cây cọ dầu. Để đánh giá tiềm năng của chủng vi khuẩn tác giả đã nghiên cứu sâu hơn về bộ gen chủng CP22 bằng phương pháp giải trình tự, chú thích và phân tích bộ gen của vi khuẩn. Kết quả thu được chủng CP22 có kích thước 5826203 bp với hàm lượng G+C là 70,84%. Phân tích chi tiết bộ gen về sự phân hủy lignocellulose cho thấy tổng cộng 60 gen bao gồm 47 gen liên quan đến sự thủy phân glycoside và 16 gen liên quan đến phân cắt liên kết este trong cacbohydrat. Đặc biệt, 20 gen mã hóa cho cellulase (8 endoglucanase, 3 exoglucanase và 9 β-glucosidase) và 40 gen mã hóa cho hemicellulose (15 endo- 1,4-β-xylanase, 3β-xylosidase, 3α-arabinofuranosidase, 10 acetyl xylan esterase, 6 polysacharide deacetylase, 1 β-mannanase, 1β-mannosidase và 1 α-galactosidase). Ba mươi gen mã hóa modun liên kết carbohydrate (CBM). Các CBM này được tìm thấy trong 27 gen phân giải cellulose và hemicellulose, cho thấy vai trò tiềm năng của chúng trong việc tăng cường liên kết với cơ chất của các enzym. Các dữ liệu phân tích bộ gen của chủng CP22 đã tạo điều kiện thuận lợi cho việc khám phá chủng vi khuẩn có khả năng phân giải lignocellulose góp phần ứng dụng công nghệ sinh học trong thực tiễn [119].
Khả năng sinh tổng hợp endoglucanase, exoglucanase, -glucosidase, xylanase của MP1 đã được chứng minh ở các nghiên cứu phần trên.
Nhận thấy chủng chủng MP1 có hệ enzym phong phú, có tiềm năng lớn trong phân giải các vật liệu giàu cellulose vi vậy chúng tôi tiếp tục thu nhận enzym thô của chủng MP1 để thực hiện các nghiên cứu khảo sát khả năng thủy phân lignocelluloses của chủng MP1.
3.6. Khảo sát khả năng thủy phân cellulose của rơm sử dụng cellulase thu nhận từ MP1
3.6.1. Xác định thành phần nguyên liệu rơm
Rơm được lấy từ ruộng, ngâm sạch, sấy khô, xử lý, nghiền nhỏ và xác định thành phần hóa học
Thành phần hóa học của rơm gồm có cellulose, hemicellulose, lignin và tro. Trước tiền xử lý các thành phần của rơm xác định được cellulose 38,4%, hemicellulose 26,7% và lignin 13,8% , Tuy nhiên sau tiền xử lý các thành phần này thay đổii đáng kể (bảng 3.27)
Bảng 3.18 Một số thành phần hóa học của rơm trước và sau tiền xử lý bằng kiềm (% chất khô nguyên liệu)
Cellulose | Hemicellulose | Lignin | |
Rơm chưa xử lý | 38,4 ± 1,71 | 26,7± 1,3 | 13,8± 0,5 |
Rơm đã tiền xử lý bằng NaOH | 55,3 ± 1,43 | 12,8± 1,7 | 11,6± 1,3 |
So sánh với các kết quả về thành phần của rơm do Bernard và cộng sự (2020) đã công bố cho thấy tương tự với kết quả chúng tôi xác định [120]. Sau khi tiền xử lý rơm bằng phương pháp kiềm hàm lượng cellulose trong rơm tăng từ 38,4% lên tới 55,3% nhờ quá trình tiền xử lý đã loại bỏ đi một lượng đáng kể hemicellulose và lignin. Với rơm sau tiền xử lý cellulose trở nên thành phần chính trong nguyên liệu chiếm 55,3% [61].
3.6.2. Ảnh hưởng của tiền xử lý cơ chất tới hiệu suất quá trình thủy phân
Rơm trước và sau tiền xử lý bằng kiềm được sử dụng làm nguyên liệu cho quá trình thủy phân sử dụng enzym thô thu nhận từ chủng vi khuẩn Cellulosimicrobium sp. MP1. Quá trình đường hóa được thực hiện ở nhiệt độ 50°C với tỷ lệ cơ chất/chất lỏng 1/100 (g/ml), nồng độ enzym 150 U/100ml. Xác định hàm lượng đường khử tạo thành sau 6,18 và 24 giờ thủy phân.