29
nguồn cung cấp một phần nhu cầu chất đạm cho người và gia súc, góp phần tạo được sự cân bằng dinh dưỡng trong khẩu phần ăn.
Đậu tương được coi là cây lấy hạt quan trọng hàng đầu trên thế giới chỉ sau lúa mì, lúa nước và ngô. Các sản phẩm từ đậu tương được sử dụng rộng rãi ở mọi nơi trên thế giới. Hiện nay, đậu tương là nguyên liệu cho nhiều ngành công nghiệp chế biến như: thực phẩm, cao su nhân tạo, sơn, mực in, xà phòng, chất dẻo, dầu bôi trơn… Trên thế giới mức sản xuất đậu tương đã tăng lên nhiều nhất trong số những cây trồng chính để đáp ứng các nhu cầu ngày càng tăng [92][96][97][98]. Ở Việt Nam, nhu cầu về đậu tương luôn ở mức cao, do đó trong "Kế hoạch phát triển nông nghiệp và nông thôn giai đoạn 2016 - 2020" của Chính phủ đã đặt ra mục tiêu đến năm 2020 đạt qui mô sản xuất với diện tích canh tác 166 nghìn ha, tổng sản lượng 265 nghìn tấn. Tuy nhiên, diện tích canh tác và tổng sản lượng đậu tương đến năm 2020 chỉ đạt 41,6 nghìn ha và 65,4 nghìn tấn (FAO). Theo số liệu của tổng cục hải quan năm 2017, lượng nhập khẩu đậu tương của cả nước khoảng 918,72 nghìn tấn, trị giá 391,93 triệu USD từ các thị trường như Mỹ, Brazil và Canada, trong đó Mỹ là thị trường cung cấp chính. Như vậy, qui mô sản xuất đậu tương trong nước thấp hơn nhiều so với nhu cầu cũng như so với mục tiêu đặt ra. Nước ta có đủ điều kiện để canh tác cây đậu tương phục vụ nhu cầu trong nước. Do đó, để có thể phát triển sản xuất, tăng sản lượng đậu tương, ngoài mở rộng thêm diện tích trong cơ cấu luân canh, cải thiện kỹ thuật canh tác và các yếu tố thương mại khác, thì công tác tạo giống mới cũng là một nhiệm vụ hàng đầu.
1.3.2. Nghiên cứu phát sinh hình thái ở đậu tương Nghiên cứu phát sinh hình thái chồi và rễ
Nhiều nghiên cứu phát sinh chồi bất định ở đậu tương đã được thực hiện trên các giống khác nhau. Các nghiên cứu cho thấy nhiều loại mẫu có thể được sử dụng để cảm ứng tạo chồi như đốt lá mầm, một nửa hạt, trụ trên và dưới lá mầm… Trong đó mẫu mô phân sinh ở vùng đốt lá mầm được ghi nhận có khả năng tạo chồi tốt nhất trên hầu hết các giống. Mức độ tạo chồi của mẫu phụ thuộc vào một số yếu tố như môi trường (khoáng), chất điều hòa sinh trưởng, nồng độ chất điều hòa, giống đậu tương... Để tạo chồi bất định, mẫu cần được cảm ứng bởi một hoặc nhiều chất điều hòa sinh trưởng, chủ yếu là cytokinin. Các nghiên cứu chủ yếu tập trung khảo
30
sát hiệu quả của việc tạo chồi bằng cách sử dụng các chất điều hòa sinh trưởng khác nhau, sử dụng đơn lẻ hoặc kết hợp, đồng thời thay đổi nồng độ chất điều hòa sinh trưởng để tìm cách thức cảm ứng chồi tốt nhất. Hầu hết các loại cytokinin gồm BA, TDZ, kinetin, zeatin đều đã được sử dụng. Ngoài ra, các nghiên cứu tạo rễ cho chồi in vitro để tạo cây con hoàn chỉnh cũng đã được thực hiện trên môi trường có hoặc không bổ sung auxin. Sau đây là những nghiên cứu cụ thể.
Nghiên cứu tạo chồi in vitro đầu tiên ở đậu tương được thực hiện bởi Cheng và cộng sự (1980), sử dụng BA (5 - 50 µM) để cảm ứng chồi từ đốt lá mầm (giống Amsoy và Dare, Mỹ). Nghiên cứu đã mô tả cách thu nhận mẫu đốt lá mầm cũng như chứng tỏ với sự cảm ứng của BA, mẫu đốt lá mầm có thể tạo nhiều chồi bất định [99]. Các nghiên cứu tạo chồi in vitro về sau hầu hết đều sử dụng mẫu đốt lá mầm thu nhận theo những mô tả này. Tác giả cũng ghi nhận khả năng hình thành rễ của chồi trên môi trường MS không bổ sung chất điều hòa sinh trưởng, các cây in vitro này sau đó có thể thích nghi và phát triển trong điều kiện vườn ươm.
Có thể bạn quan tâm!
-

 Đối Tượng, Phạm Vi Và Nội Dung Nghiên Cứu Của Đề Tài
Đối Tượng, Phạm Vi Và Nội Dung Nghiên Cứu Của Đề Tài -
![Vị Trí Của Phân Tử Astaxanthin Trong Màng Tế Bào [40]](https://tailieuthamkhao.com/uploads/2022/12/28/nghien-cuu-tao-cay-dau-tuong-glycine-max-l-bien-doi-gen-co-kha-nang-tong-4-1-120x90.jpg) Vị Trí Của Phân Tử Astaxanthin Trong Màng Tế Bào [40]
Vị Trí Của Phân Tử Astaxanthin Trong Màng Tế Bào [40] -
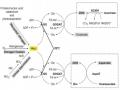 Sự Ức Chế Của Phosphinothricin Lên Hoạt Động Của Enzyme Glutamine Synthetase (Gs)
Sự Ức Chế Của Phosphinothricin Lên Hoạt Động Của Enzyme Glutamine Synthetase (Gs) -
 Chuyển Gen Vào Đậu Tương Bằng Phương Pháp Bắn Gen
Chuyển Gen Vào Đậu Tương Bằng Phương Pháp Bắn Gen -
 Kim Châm Để Tạo Vết Thương Cho Mẫu Chuyển Gen
Kim Châm Để Tạo Vết Thương Cho Mẫu Chuyển Gen -
 Ảnh Hưởng Của Ppt Lên Khả Năng Tái Sinh Của Đốt Lá Mầm Và Một Nửa Hạt
Ảnh Hưởng Của Ppt Lên Khả Năng Tái Sinh Của Đốt Lá Mầm Và Một Nửa Hạt
Xem toàn bộ 176 trang tài liệu này.
Franklin và cộng sự (2004) nghiên cứu khả năng cảm ứng tạo chồi của BA và TDZ trên đốt lá mầm một số giống đậu tương ở Mỹ (PNP, Dekalb, Sandusky, CNRR 279, CB 277). Trên giống PNP, kết quả cho thấy BA và TDZ khi được sử dụng riêng lẻ hay kết hợp đều có khả năng cảm ứng tạo chồi khá tốt, với nồng độ BA cố định 13,3 µM cho tỉ lệ mẫu tạo chồi 50%; 3,2 chồi/mẫu và nồng độ TDZ (4,45 – 22,71 µM) tối ưu 4,54 µM cho hiệu quả tạo chồi 68%, 7,3 chồi/mẫu; sự kết hợp 4,54 µM TDZ + 13,3 µM BA dẫn đến tăng khả năng cảm ứng chồi so với sử dụng riêng lẻ từng chất (84%; 19,2 chồi/mẫu). Khi áp dụng môi trường tối ưu với các giống khác, tác giả nhận thấy có sự thay đổi về khả năng tái sinh tuy nhiên hiệu quả tái sinh vẫn khá cao (>69%) [100]. Ngoài ra, nghiên cứu cũng cho thấy khi bổ sung NAA vào môi trường đã giúp tăng đáng kể khả năng tạo rễ của chồi in vitro, với tỉ lệ chồi tạo rễ đạt cao nhất 88,3% ở nồng độ 2,69 µM so với đối chứng không bổ sung NAA là 12,5%.
Nghiên cứu của Janani và Kumari (2013) trên giống JS335 Ấn Độ cũng sử dụng TDZ và BA để cảm ứng tạo chồi từ đốt lá mầm, trong đó cả BA và TDZ đều được khảo sát các nồng độ khác nhau (5-25 µM) để tìm nồng độ tối ưu. Kết quả ghi nhận cho thấy cả BA và TDZ đều có khả năng cảm ứng tạo chồi tốt, ở nồng độ tối

31
ưu TDZ (15 µM) cảm ứng tạo chồi tốt hơn so với nồng độ BA tối ưu (15 µM) (80%, 8,33 chồi/mẫu) so với (53,3%, 4,7 chồi/mẫu). Tăng nồng độ quá mức tối ưu, cả TDZ và BA đều dẫn đến giảm khả năng cảm ứng chồi [101]. Như vậy, nghiên cứu trên các giống đậu tương Ấn Độ và Mỹ đều cho thấy TDZ và BA có khả năng cảm ứng tạo chồi tốt trên mẫu đốt lá mầm trong đó TDZ có phần hiệu quả hơn, xét về tỉ lệ mẫu tạo chồi và số chồi/mẫu [100][101]. Ngoài ra có thể thấy, nồng độ tối ưu của TDZ thay đổi khá lớn giữa giống của Mỹ và Ấn Độ, trong khi nồng độ tối ưu giống PNP (Mỹ) chỉ 4,45 µM, giống JS335 (Ấn Độ) cần 15 µM. Đối với BA, nồng độ tối ưu của giống Ấn Độ khá gần với nồng độ sử dụng với giống Mỹ (13,3 và 15 µM). Chồi tạo thành (giống JS335 Ấn Độ) được cảm ứng tạo rễ trên môi trường MS bổ sung IBA ở các nồng độ khác nhau từ 5 đến 25 µM. Kết quả ghi nhận, trên môi trường bổ sung 20 µM IBA chồi có khả năng tạo rễ tốt nhất với 41,6 rễ/chồi, khi tăng lên 25 µM IBA làm giảm khả năng tạo rễ, còn 36,3 rễ/chồi.
Ma và Wu (2008) khảo sát khả năng tạo chồi từ đốt lá mầm nguyên (giữ nguyên hai lá mầm, không chẻ đôi) của một số giống đậu tương Trung Quốc (Jilin 35, Dongnong 42, Hefeng 25, Hefeng 41) cho thấy 3 mg/l BA có khả năng cảm ứng tạo chồi cao với tỉ lệ mẫu tạo chồi trung bình 97,2%, 13 chồi/mẫu. Tỉ lệ mẫu tạo chồi không có nhiều khác biệt giữa các giống, tuy nhiên số chồi/mẫu thay đổi từ 7-18. Ngoài ra, nghiên cứu ghi nhận sự kết hợp nồng độ tối ưu BA 3 mg/l với hàm lượng nhỏ kinetin 0,5 mg/l và IBA 0,2 mg/l giúp tăng số chồi nhận được, tối ưu ở giống Hengfeng 25 với 30 chồi/mẫu [102].
Zia và cộng sự (2010) nghiên cứu tái sinh chồi từ đốt lá mầm 2 giống đậu tương NARC-4 và NARC-7 (Parkistan) sử dụng riêng lẻ từng chất BAP, Kinetin và Zeatin. Kết quả cho thấy cả ba chất đều có khả năng cảm ứng tạo đa chồi trong đó 1 mg/l BAP cảm ứng tạo chồi tốt nhất ở cả hai giống NARC-4 (88% mẫu tạo chồi; 7,3 chồi/mẫu) và NARC-7 (82% mẫu tạo chồi; 6,4 chồi/mẫu). Khi tăng nồng độ lên 2 mg/l khả năng cảm ứng tạo chồi của BAP, Kinetin và Zeatin đều giảm ở cả hai giống. Ngoài ra, IBA được ghi nhận giúp tăng đáng kể khả năng tạo rễ cho chồi in vitro với nồng độ tối ưu là 1 mg/l cả giống NARC-4 (76,3% chồi tạo rễ; 4,7 rễ/chồi) và NARC-7 (55,3% chồi tạo rễ; 2,2 rễ/chồi) so với đối chứng chỉ khoảng 30% chồi
32
tạo rễ, tăng nồng độ IBA dẫn đến giảm khả năng tạo rễ ở cả tỉ lệ chồi tạo rễ và số rễ/chồi của hai giống [103].
Soto và cộng sự (2013) sử dụng BAP (0,5-6 mg/l) trên giống Incasoy-36 của Cuba, nhận thấy tỉ lệ tạo chồi tối ưu với BAP 1,5 mg/l (96,8%, 4,3 chồi/mẫu). Tăng nồng độ BAP trên 1,5 mg/l làm giảm khả năng tạo chồi, đồng thời ở nồng độ quá cao 6 mg/l, ghi nhận hiện tượng tạo nhiều chồi nhỏ bất thường. Nồng độ BAP từ 3 mg/l trở lên cũng dẫn đến tạo nhiều mô sẹo ở vùng trụ dưới lá mầm, ảnh hưởng xấu tới sự tạo và phát triển chồi [104]. Trên giống đậu tương này, các tác giả ghi nhận chồi in vitro có khả năng tạo rễ tốt, với tỉ lệ chồi tạo rễ 97%, trên môi trường MS không bổ sung chất điều hòa sinh trưởng.
Mangena và cộng sự (2015) nghiên cứu khả năng cảm ứng tạo chồi từ đốt lá mầm của BA và Kinetin trên giống giống LS 677, Nam Phi. Kết quả cho thấy 2 mg/l BA cho hiệu quả cảm ứng chồi tốt nhất với tỉ lệ mẫu tạo chồi 56,7%, 5,27 chồi/mẫu. Khi tăng nồng độ BA lên 4 mg/l dẫn đến giảm khả năng cảm ứng tạo chồi đồng thời kích thích tạo nhiều mô sẹo. Các tác giả cũng ghi nhận sự kết hợp của BA ở nồng độ tối ưu (2 mg/l) và 1 mg/l Kinetin làm giảm khả năng cảm ứng chồi với tỉ lệ mẫu tạo chồi giảm còn 13,3%, 2 chồi/mẫu. Ngoài ra, nghiên cứu cũng cho thấy mẫu đốt lá mầm không bị tách đôi cho hiệu quả tạo chồi cao hơn so với mẫu đốt lá mầm thông thường [105]. Với khảo sát tạo rễ, chồi in vitro được ghi nhận có khả năng hình thành rễ tốt trên môi trường MS không bổ sung chất điều hòa sinh trưởng, cây con tạo thành cũng có thể thích ứng, phát triển trong vườn ươm, với tỉ lệ cây sống khoảng 70%.
Raza và cộng sự (2017) sử dụng nhiều loại mẫu khác nhau gồm trụ hạ diệp, trụ hạ diệp tách đôi và đốt lá mầm của 9 giống đậu tương (Australia) để cảm ứng tạo chồi với 1,67 mg/l BA. Kết quả cho thấy khả năng cảm ứng tạo chồi ở các giống khác nhau với tỉ lệ tạo chồi và số chồi/mẫu thay đổi: trụ hạ diệp (50 - 100%; 2,7 - 4,2 chồi/mẫu); trụ hạ diệp tách đôi (60 - 87%; 4,2 - 10 chồi/mẫu); đốt lá mầm (75 - 100%, 2,6 - 10,5 chồi/mẫu). Như vậy, mặc dù trụ hạ diệp cũng có thể tạo chồi khá tốt ở một số giống khi cảm ứng bởi BA, nhưng nhìn chung đốt lá mầm vẫn có hiệu quả tốt hơn ở hầu hết giống [106]. Để tạo rễ cho chồi in vitro, các tác giả sử dụng
33
IBA ở nồng độ cố định 1 mg/l bổ sung vào môi trường MS, kết quả ghi nhận cho thấy hầu hết chồi đều có khả năng tạo rễ tốt.
Các nghiên cứu trên cho thấy BA, một cách xuyên suốt có khả năng cảm ứng tạo chồi tốt trên nhiều giống đậu tương. Nồng độ tối ưu thay đổi khác nhau phụ thuộc vào giống (từ 1 đến 3 mg/l), khi vượt quá nồng độ tối ưu, sẽ dẫn đến làm giảm khả năng cảm ứng chồi. Giữa các giống khác nhau khả năng cảm ứng chồi tối ưu cũng thay đổi đáng kể, từ khoảng 55% (3 chồi/mẫu) - 100% (10 chồi/mẫu). Với khả năng tạo rễ của chồi in vitro, mặc dù ở một số giống, chồi có thể tạo rễ không cần bổ sung auxin ngoại sinh, tuy nhiên các nghiên cứu cho thấy, khi được bổ sung vào môi trường nuôi cấy, auxin (IBA hoặc NAA) đều giúp tăng đáng kể hiệu quả tạo rễ ở cả tỉ lệ mẫu tạo rễ và số rễ/chồi.
Các nghiên cứu tái sinh tạo chồi bất định từ đốt lá mầm của các giống đậu tương trong nước có kết quả khá giống với nghiên cứu nước ngoài. Hầu hết các nghiên cứu đều dùng BA để cảm ứng tạo chồi, nồng độ BA tối ưu đạt được cũng thay đổi từ 1-2 mg/l với khả năng cảm ứng chồi tối ưu của các giống thay đổi từ khoảng 84% (2 chồi/mẫu) - 98% (5,5 chồi/mẫu). Chồi in vitro được cảm ứng tạo rễ trên môi trường MS bổ sung IBA với nồng độ tối ưu khoảng 1 mg/l trong các nghiên cứu khác nhau.
Cụ thể các nghiên cứu như sau:
Nghiên cứu tái sinh đậu tương cũng đã được một số tác giả trong nước thực hiện. Nguyễn Thị Thu Hường và cộng sự (2009) nghiên cứu khả năng cảm ứng tạo chồi của BA trên đốt lá mầm 2 giống đậu tương ĐT12 và ĐT84 cho thấy 1,5 mg/l BA có khả năng cảm ứng tạo chồi tốt nhất trên cả hai giống ĐT 12 (91,2% mẫu tạo chồi; 2,3 chồi/mẫu) và ĐT 84 (92,1% mẫu tạo chồi; 2,2 chồi/mẫu). Khi tăng nồng độ BA lên 1,67 mg/l đều dẫn đến giảm đáng kể hiệu quả cảm ứng chồi trên cả hai giống. Ngoài ra, nghiên cứu cũng cho thấy việc tạo vết thương cho vùng đốt lá mầm là cần thiết để cảm ứng tạo đa chồi. Nghiên cứu cũng ghi nhận 1 mg/l IBA có khả năng cảm ứng tạo rễ tốt nhất cho chồi ở cả 2 giống ĐT 12 (98% chồi tạo rễ) và ĐT 84 (97% chồi tạo rễ). Tăng nồng độ IBA lên 1,5 và 2 mg/l đều làm giảm khả năng tạo rễ của chồi đáng kể [107].
34
Nguyễn Tiến Dũng và Ngô Xuân Bình (2011) nghiên cứu ảnh hưởng của BA và Kinetin đến khả năng tái sinh chồi từ đốt lá mầm của 10 giống đậu tương Việt Nam đã nhận thấy BA có khả năng cảm ứng tạo đa chồi tốt hơn so với Kinetin. Nồng độ tối ưu của BA thay đổi tùy giống và nằm trong khoảng 1 - 2 mg/l với tỉ lệ mẫu tạo chồi từ 84 - 98%, số chồi/ mẫu từ 2,1 - 3,57 [108].
Phan Lê Tư và cộng sự (2018) sử dụng 1 mg/l BA cảm ứng tạo đa chồi từ đốt lá mầm của 6 giống đậu tương Việt Nam cho thấy sử dụng 1 mg/l có khả năng kích thích tạo đa chồi khá tốt trên hầu hết các giống với tỉ lệ mẫu tạo chồi từ 78,7 - 94,7%, số chồi/mẫu từ 2,86 - 5,49 [109]. Chồi in vitro trong nghiên cứu có thể tạo rễ tốt trong môi trường bổ sung IBA 1 mg/l.
Ngoài ra, nghiên cứu chuyển gen thông qua vi khuẩn A. tumefaciens trên đậu tương phần lớn đều sử dụng mẫu đốt lá mầm làm vật liệu chuyển gen với BA là chất cảm ứng tạo chồi. Nồng độ BA thay đổi tùy giống đậu tương, thường biến động trong khoảng 1 - 2 mg/l [110][111][112]. Để cảm ứng rễ chồi chuyển gen, hầu hết nghiên cứu sử dụng IBA với nồng độ khoảng 1 mg/l, trong môi trường hoặc nhúng trước khi chuyển vào môi trường nuôi cấy.
Như vậy, nhiều nghiên cứu cảm ứng tạo chồi bất định từ đốt lá mầm trên các giống đậu tương khác nhau trong và ngoài nước đã cho thấy, trong các loại cytokinin được sử dụng khảo sát TDZ và BA có khả năng cảm ứng tạo chồi tốt nhất, khả năng này thể hiện trên hầu hết các giống khác nhau, tuy nồng độ tối ưu thay đổi tùy giống. Các nghiên cứu của Franklin và cộng sự (2004), Janani và Kumari (2013) cho thấy TDZ có phần hiệu quả hơn so với BA trong việc cảm ứng tạo chồi, tuy nhiên các nghiên cứu này chưa đánh giá mức độ đồng đều, khả năng phát triển của các chồi này thành cây hoàn chỉnh [100] [101]. Một số nghiên cứu cho thấy TDZ dễ tạo các dạng tái sinh bất thường [113][114]. Hơn nữa, theo ghi nhận, hầu hết các nghiên cứu chuyển gen thông qua vi khuẩn A. tumefaciens dùng mẫu đốt lá mầm, một nửa hạt tạo được cây đậu tương chuyển gen hữu thụ đều sử dụng BA để cảm ứng chồi [110][111][112], điều này có thể do, BA vừa có khả năng cảm ứng chồi tốt trên hầu hết giống, các chồi này cũng có khả năng phát triển bình thường và tạo cây chuyển gen hữu thụ.
35
Tóm lại, dựa trên các nghiên cứu cảm ứng chồi, tạo tạo rễ cho chồi in vitro của nhiều giống đậu tương, cũng như các nghiên cứu chuyển gen vào đậu tương, BA và IBA lần lượt được ghi nhận là loại cytokinin, auxin thích hợp để cảm ứng chồi, rễ trong giai đoạn nuôi cấy mô, ứng dụng cho nghiên cứu chuyển gen trên nhiều giống khác nhau, tuy nhiên vẫn cần khảo sát để tìm nồng độ thích hợp nhất giúp tạo cây con hoàn chỉnh.
Nghiên cứu tạo phôi soma
Christianson và cộng sự (1983) lần đầu tiên mô tả sự tái sinh thông qua tạo phôi soma của đậu tương [115]. Sau đó, Lazzeri và cộng sự (1985) đã thành công khi sử dụng lá mầm chưa trưởng thành để cảm ứng tạo phôi soma [116]. Từ đó, hầu hết các nghiên cứu sau này đã sử dụng lá mầm chưa trưởng thành để tạo phôi soma [117][118]
Nhiều loại auxin đã được sử dụng để cảm ứng tạo phôi soma ở đậu tương, trong đó 2,4-D cho thấy hiệu quả trong nhiều nghiên cứu [119]. Sự phát sinh phôi soma từ lá mầm chưa trưởng thành phụ thuộc nhiều vào giống [27][120].
1.3.3. Nghiên cứu biến đổi gen đậu tương
Hai phương pháp được sử dụng phổ biến nhất để chuyển gen vào đậu tương là phương pháp chuyển gen gián tiếp thông qua vi khuẩn A. tumefaciens và phương pháp bắn gen. Mỗi phương pháp đều có những ưu nhược điểm riêng và được lựa chọn sử dụng tùy thuộc vào điều kiện trang thiết bị, giống, chủng vi khuẩn, thực vật…
1.3.3.1. Chuyển gen vào đậu tương sử dụng vi khuẩn A. tumefaciens
Khả năng xâm nhiễm tự nhiên của vi khuẩn A. tumefaciens trên đậu tương thấp hơn nhiều so với các cây hai lá mầm khác như thuốc lá… Do đó, để biến nạp gen vào đậu tương cần nhiều cải tiến để nâng cao hiệu quả chuyển gen. Mặc dù, nhiều nghiên cứu đã góp phần tăng khả năng chuyển gen, tuy nhiên đến nay đậu tương vẫn được xem là một đối tượng khó chuyển gen, hiệu quả chuyển gen phụ thuộc nhiều vào giống và kỹ năng của người thực hiện. Nhiều loại mẫu khác nhau có thể được dùng để chuyển gen như đốt lá mầm, một nửa hạt, trụ hạ diệp, lá mầm non, đỉnh chồi… Trong đó, đốt lá mầm và một nửa hạt được sử dụng phổ biến nhất do có hiệu quả tái sinh chồi cao trên hầu hết giống với điều kiện tái sinh tương đối
36
đơn giản, trong môi trường MS, thường bổ sung 1-3 mg/l BA. Chuyển gen thông qua sự hình thành phôi gặp hạn chế ở khả năng nuôi cấy tạo phôi kém ở nhiều giống đậu tương. Các nghiên cứu tăng hiệu quả chuyển gen vào đậu tương thường tập trung giải quyết các vấn đề như: hệ thống chọn lọc cây chuyển gen, bổ sung các chất chống oxy hóa để hạn chế tình trạng hóa nâu của mẫu, tối ưu các phương pháp tạo vết thương và các thông số cơ bản trong qui trình chuyển gen: mật độ vi khuẩn, thời gian đồng nuôi cấy,... Sau đây là các nghiên cứu nhằm tối ưu hiệu quả chuyển gen trên nhiều giống đậu tương khác nhau trên thế giới và ở Việt Nam.
Sử dụng hệ thống chọn lọc phù hợp gồm gen và chất chọn lọc đóng vai trò quan trọng đến khả năng chuyển gen thành công vào thực vật. Ở đậu tương, một số hệ thống chọn lọc đã được sử dụng, đánh giá trong việc chọn lọc cây chuyển gen. Hinchee và cộng sự (1988) đã lần đầu tiên tạo thành công cây đậu tương chuyển gen hữu thụ, sử dụng đốt lá mầm và vi khuẩn A. tumefaciens [121]. Các tác giả sử dụng gen chọn lọc neomycin phosphotransferase II (npt II) với chất chọn lọc là kanamycin 200 - 300 mg/l. Hệ thống này cho thấy nhược điểm khi có xác suất tạo thể khảm và khả năng “escape” cao dẫn đến sự tồn tại của nhiều chồi không chuyển gen. Sau quá trình chọn lọc, nhận được 128 cây tuy nhiên chỉ 8 cây (6%) chuyển gen. Ngoài ra, khả năng chọn lọc thành công cũng phụ thuộc lớn vào giống đậu tương được sử dụng. Nghiên cứu của Rani và cộng sự (2012) cũng cho thấy chỉ 20 cây chuyển gen trong 91 cây kháng với kanamycin [122]. Di và cộng sự (1996) cũng ghi nhận khả năng chọn lọc không tốt của kanamycin [123].
Khắc phục các nhược điểm của hệ thống này, Zhang và cộng sự (1999) lần đầu tiên đã sử dụng gen chọn lọc bar, với chất chọn lọc là thuốc diệt cỏ glufosinate trên đậu tương [112]. Các tác giả sử dụng đốt lá mầm từ cây nảy mầm 5 ngày tuổi. Thử nghiệm 2 hệ thống chọn lọc với nồng độ thay đổi ở giai đoạn tạo chồi và kéo dài chồi (3,3 mg/l; 1,7 mg/l và 5 mg/l; 2 mg/l). Kết quả cho thấy chọn lọc với nồng độ 5 mg/l giai đoạn đầu và 2 mg/l giai đoạn sau cho kết quả tốt nhất với hiệu quả chuyển gen đạt được là 3%; với hệ thống chọn lọc ở nồng độ glufosinate thấp hơn, hiệu quả chuyển gen đạt được chỉ 0,5%. Hệ thống này cho khả năng chuyển gen thành công trên nhiều giống, ít tạo thể khảm, cây chuyển gen ổn định về di truyền. Tuy nhiên hiệu quả chuyển gen vẫn chưa được nâng cao. Zeng và cộng sự (2004)