93
gen biến nạp do đó chưa được chứng minh. Điều này chỉ có thể được xác định thông qua sự hiện diện và hàm lượng chất mục tiêu astaxanthin trong hạt đậu chuyển gen như đã đặt ra trong mục tiêu nghiên cứu.
3.11.2. Chuyển gen bằng vi khuẩn A. tumefaciens sử dụng dao mổ kết hợp sóng siêu âm, thấm chân không tạo vết thương mẫu
3.11.2.1. Chuyển gen tạo astaxanthin vào đậu tương
Các khảo sát trên đã tối ưu phương pháp sử dụng sóng siêu âm, thấm hút chân không để xử lý, tạo vết thương mẫu giúp nâng cao hiệu quả chuyển gen gus vào đốt lá mầm đậu tương. Các thông số này được áp dụng trong nghiên cứu chuyển gen tạo cây đậu tương có khả năng tổng hợp astaxanthin chuyên biệt ở hạt. Ngoài ra, bên cạnh đốt lá mầm, mẫu một nửa hạt cũng được sử dụng làm vật liệu chuyển gen nhằm so sánh hiệu quả tiếp nhận gen so với mẫu đốt lá mầm.
Chọn lọc, tái sinh, tạo cây kháng PPT
Nghiên cứu chuyển gen thông qua vi khuẩn A. tumefaciens sử dụng mẫu đốt lá mầm và một nửa hạt, trong đó có sự hỗ trợ của sóng siêu âm, thấm chân không đã tạo được một số dòng kháng PPT. Cụ thể quá trình từ sau khi đồng nuôi cấy đến khi chuyển cây ra vườn ươm như sau: Sau khi đồng nuôi cấy 5 ngày, hầu hết đốt lá mầm và một nửa hạt vẫn giữ được trạng thái tốt: mô chắc, không bị nhiễm quá nhiều khuẩn. Mẫu sau đó được rửa và thấm ráo để loại bớt vi khuẩn và cấy lên môi trường tái sinh chọn lọc SI 6 mg/l PPT. Sau 2 tuần đầu, chất chọn lọc đã gây chết, ức chế tái sinh ở vùng mô phân sinh của nhiều mẫu, chuyển sang màu nâu đen, trong khi đó một số mẫu bắt đầu hình thành các cụm chồi nhỏ. Cấy chuyền mẫu sang môi trường SI mới, một số cụm chồi phát triển, tuy nhiên ở cuối giai đoạn tái sinh chọn lọc, nhiều cụm chồi không vươn cao, lá vàng, đen, cả cụm chồi chết dần. Hiện tượng cụm chồi có thể hình thành, rồi chết dần trên môi trường chọn lọc có thể do những mô lân cận trên lá mầm đã được biến nạp gen, đã chuyển hóa PPT ở vùng mô phân sinh tiếp xúc giúp các mô hình thành được chồi và sinh trưởng được trong giai đoạn đầu khi cụm chồi chưa phát triển lớn và chưa bị tách rời. Ngoài ra, các hợp chất thiol trong môi trường đồng nuôi cấy cũng được ghi nhận tạo tính kháng lại chất chọn lọc PPT nên gia tăng khả năng tạo chồi cho mẫu không chuyển gen dưới tác động của PPT [180]. Sau 4 tuần trên môi trường chọn lọc SI, chỉ một số ít
cụm chồi phát triển, được tách rời khỏi lá mầm và cấy chuyền sang môi trường tăng trưởng thân SE bổ sung 4 mg/l PPT.
Có thể bạn quan tâm!
-
 Sự Hình Thành Chồi Của Đốt Lá Mầm Giống Mtđ 176 Ở Các Nồng Độ Ba Khác Nhau Sau 3 Tuần Nuôi Cấy.
Sự Hình Thành Chồi Của Đốt Lá Mầm Giống Mtđ 176 Ở Các Nồng Độ Ba Khác Nhau Sau 3 Tuần Nuôi Cấy. -
 Sự Tái Sinh Của Đốt Lá Mầm Giống Mtđ 176 Trên Môi Trường Có Ppt.
Sự Tái Sinh Của Đốt Lá Mầm Giống Mtđ 176 Trên Môi Trường Có Ppt. -
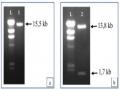 Tạo Dòng Agrobacterium Tumefaciens Eha 105 Chứa Plasmid Pitb-Ast Ly Trích, Kiểm Tra Plasmid Pitb-Ast Từ E. Coli
Tạo Dòng Agrobacterium Tumefaciens Eha 105 Chứa Plasmid Pitb-Ast Ly Trích, Kiểm Tra Plasmid Pitb-Ast Từ E. Coli -
 Dòng Chuyển Gen D2 (A) Và D8 (B) Phát Triển Ngoài Vườn Ươm
Dòng Chuyển Gen D2 (A) Và D8 (B) Phát Triển Ngoài Vườn Ươm -
 So Sánh Kiểu Hình Cây Chuyển Gen Và Cây Đối Chứng.
So Sánh Kiểu Hình Cây Chuyển Gen Và Cây Đối Chứng. -
 R.p. Mcquinn, B. Wong, J.j. Giovannoni, Atpds Overexpression In Tomato: Exposing Unique Patterns Of Carotenoid Self‐Regulation And An Alternative Strategy For The Enhancement Of Fruit
R.p. Mcquinn, B. Wong, J.j. Giovannoni, Atpds Overexpression In Tomato: Exposing Unique Patterns Of Carotenoid Self‐Regulation And An Alternative Strategy For The Enhancement Of Fruit
Xem toàn bộ 176 trang tài liệu này.
Đối với hai giống HL 07-15, OMĐN 29, tất cả cụm chồi còn sót lại trên môi trường SI đều chết dần và không nhận được cây chuyển gen.
Đối với giống MTĐ 176, hầu hết các cụm chồi cũng chết dần, chỉ một số ít tiếp tục phát triển. Các chồi này khi cao khoảng 3 cm được cấy chuyền sang môi trường tạo rễ RM bổ sung 1 mg/l PPT. Sau khoảng 3 - 4 tuần bộ rễ hình thành, cây được chuyển ra trồng ngoài vườn ươm (hình 3.21). Tần số tạo dòng kháng PPT được ghi nhận ở phương pháp dùng dao mổ tạo vết thương trên đốt lá mầm là 1/150 (0,67%), kí hiệu dòng là D1; phương pháp tạo vết thương kết hợp sóng siêu âm 30s là 3/150 (2%), kí hiệu dòng là D2, D3, D4; phương pháp tạo vết thương kết hợp thấm chân không 60s là 3/150 (2%), kí hiệu dòng là D5, D6, D7; và ở phương pháp dùng mẫu một nửa hạt là 2/200 (1%), kí hiệu dòng là D8, D9.
Các kết quả trên cho thấy sự kết hợp sóng siêu âm hoặc thấm chân không trong quá trình tạo vết thương cho mẫu đốt lá mầm đã giúp tăng đáng kể hiệu quả chuyển gen. Điều này phù hợp với kết quả khảo sát chuyển gen gus cũng như nhiều nghiên cứu chuyển gen khác thông qua vi khuẩn A. tumefaciens [131][133][134][173][176]. Ngoài ra, chuyển gen vào mẫu một nửa hạt cũng đạt được tần số khá cao, cho thấy khả năng áp dụng của loại mẫu này để chuyển gen vào đậu tương. Mặc dù hai giống đậu tương HL 07-15, OMĐN 29 có khả năng tái sinh tốt và có hình thành một số chồi kháng PPT trong các giai đoạn chọn lọc đầu, nhưng cuối cùng không nhận được cây chuyển gen. Giống MTĐ 176 đáp ứng chuyển gen tốt hơn, tất cả các phương pháp đều nhận được cây chuyển gen.
Các cây đậu tương kháng PPT phát triển khá tốt in vitro. Các cây này sau khi kiểm tra sự hiện diện của gen chuyển bằng PCR đều cho kết quả dương tính. Tuy nhiên, khi chuyển ra trồng ex vitro, trong số 9 cây chuyển gen, chỉ 5 cây D2, D3, D5, D8, D9 có khả năng thích nghi, phát triển, 4 cây không thể thích nghi và chết dần.
1 cm
a
1 cm
b
2 mm
c
5 mm
d
1 cm
e
Hình 3.21. Các bước chuyển gen và tạo cây đậu tương chuyển gen.
a:cây nảy mầm sau 7 ngày; b: mẫu đốt lá mầm đồng nuôi cấy với vi khuẩn; c: chồi hình thành sau 4 tuần trên môi trường SI bổ sung 6 mg/l PPT; d,e: chồi kéo dài trên môi trường SE bổ sung 4 mg/l PPT và tạo rễ trên môi trường RM.
3.11.2.2. Kiểm tra sự hiện diện của gen biến nạp trên các dòng kháng PPT bằng phương pháp PCR
DNA được ly trích từ lá của các dòng kháng PPT được sử dụng để thực hiện phản ứng PCR nhằm kiểm tra sự hiện diện của các gen biến nạp trong bộ gen cây, DNA của cây không chuyển gen sử dụng làm đối chứng âm (N), plasmid pITB- AST được dùng làm đối chứng dương (P).
Kiểm tra sự hiện diện của gen bar
Phản ứng PCR được thực hiện với cặp mồi đặc hiệu khuếch đại đoạn gen bar, kích thước đặc trưng là 500 bp. Kết quả điện di sản phẩm PCR (hình 3.22) như sau:
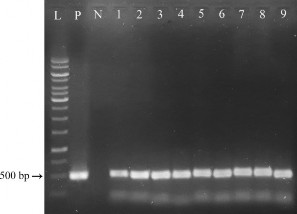
Hình 3.22. Kiểm tra sự hiện diện của gen bar ở các cây kháng PPT bằng PCR.
L: thang chuẩn 1kb (Promega); P: đối chứng dương; N: cây đối chứng; 1, 2, 3, 4, 5, 6, 7, 8, 9 tương ứng các dòng chuyển gen giả định D1, D2, D3, D4, D5, D6, D7, D8, D9
Ở mẫu đối chứng dương P (plasmid pITB-AST) và các cây chuyển gen giả định D1, D2, D3, D4, D5, D6, D7, D8, D9 đều có sự hiện diện của 1 băng DNA với kích thước khoảng 500 bp khi so sánh với thang chuẩn (phù hợp với kích thước đặc hiệu của đoạn DNA được khuếch đại là 500 bp), trong khi phản ứng PCR với mẫu đối chứng âm N (DNA của cây không chuyển gen) cho kết quả âm tính. Do đó, có thể khẳng định có sự hiện diện của gen bar trong tất cả các dòng kháng PPT. Sự biểu hiện của gen bar giúp các dòng này có thể tồn tại và phát triển trên môi trường có chất chọn lọc PPT.
Kiểm tra sự hiện diện của gen cbfd2
Phản ứng PCR được thực hiện với cặp mồi đặc hiệu khuếch đại đoạn gen cbfd2, kích thước đặc trưng là 500 bp. Kết quả điện di sản phẩm PCR (hình 3.23) như sau:
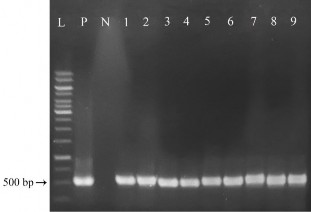
Ở mẫu đối chứng dương P (plasmid pITB-AST) và các cây chuyển gen giả định D1, D2, D3, D4, D5, D6, D7, D8, D9 đều có sự hiện diện của 1 băng DNA với kích thước khoảng 500 bp khi so sánh với thang chuẩn (phù hợp với kích thước đặc hiệu của đoạn DNA được khuếch đại là 500 bp), trong khi phản ứng PCR với mẫu đối chứng âm N (DNA của cây không chuyển gen) cho kết quả âm tính. Do đó, có thể khẳng định có sự hiện diện của gen cbfd2 trong tất cả các cây D1, D2, D3, D4, D5, D6, D7, D8, D9.
Kiểm tra sự hiện diện của gen hbfd1
Hình 3.23. Kiểm tra sự hiện diện của gen cbfd2 ở các cây kháng PPT bằng PCR.
L: thang chuẩn 1kb (Promega); P: đối chứng dương; N: cây đối chứng; 1, 2, 3, 4, 5, 6, 7, 8, 9 tương ứng các dòng chuyển gen giả định D1, D2, D3, D4, D5, D6, D7, D8, D9
Phản ứng PCR được thực hiện với cặp mồi đặc hiệu khuếch đại đoạn gen hbfd1, kích thước đặc trưng là 400 bp. Kết quả điện di sản phẩm PCR (hình 3.24) như sau:
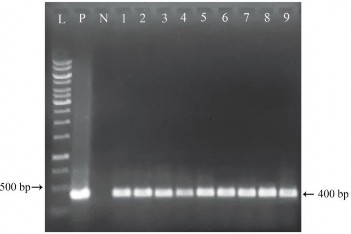
Hình 3.24. Kiểm tra sự hiện diện của gen hbfd1 ở các cây kháng PPT bằng PCR. L: thang chuẩn 1kb (Promega); P: đối chứng dương; N: cây đối chứng; 1, 2, 3, 4, 5, 6, 7, 8, 9 tương ứng các dòng chuyển gen giả định D1, D2, D3, D4, D5, D6, D7, D8, D9
Ở mẫu đối chứng dương P (plasmid pITB-AST) và các cây chuyển gen giả định D1, D2, D3, D4, D5, D6, D7, D8, D9 đều có sự hiện diện của 1 băng DNA với kích thước khoảng 400 bp khi so sánh với thang chuẩn (phù hợp với kích thước đặc hiệu của đoạn DNA được khuếch đại là 400 bp), trong khi phản ứng PCR với mẫu đối chứng âm N (DNA của cây không chuyển gen) cho kết quả âm tính. Do đó, có thể khẳng định có sự hiện diện của gen hbfd1 trong tất cả các cây D1, D2, D3, D4, D5, D6, D7, D8, D9.
Kiểm tra sự hiện diện của gen Zm-psy
Phản ứng PCR được thực hiện với cặp mồi đặc hiệu khuếch đại đoạn gen Zm-psy, kích thước đặc trưng là 446 bp. Kết quả điện di sản phẩm PCR (hình 3.25) như sau:
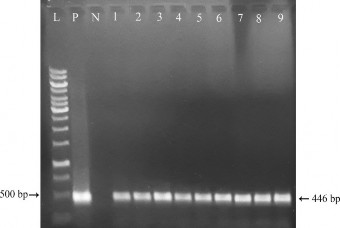
Hình 3.25. Kiểm tra sự hiện diện của gen Zm-psy ở các cây kháng PPT bằng PCR.
L: thang chuẩn 1kb (Promega); P: đối chứng dương; N: cây đối chứng; 1, 2, 3, 4, 5, 6, 7, 8, 9 tương ứng các dòng chuyển gen giả định D1, D2, D3, D4, D5, D6, D7, D8, D9
Ở mẫu đối chứng dương P (plasmid pITB-AST) và các cây chuyển gen giả định D1, D2, D3, D4, D5, D6, D7, D8, D9 đều có sự hiện diện của 1 băng DNA với kích thước khoảng 446 bp khi so sánh với thang chuẩn (phù hợp với kích thước đặc hiệu của đoạn DNA được khuếch đại là 446 bp), trong khi phản ứng PCR với mẫu đối chứng âm N (DNA của cây không chuyển gen) cho kết quả âm tính. Do đó, có thể khẳng định có sự hiện diện của gen Zm-psy trong tất cả các cây D1, D2, D3, D4, D5, D6, D7, D8, D9.
Như vậy, kết quả PCR sử dụng các cặp mồi chuyên biệt cho gen bar, hbfd1, cbfd2, Zm-psy đã giúp khẳng định các dòng kháng PPT chọn lọc được đều mang gen biến nạp. Tuy nhiên đây chỉ là kết quả bước đầu, do đó cần tiếp tục phân tích Southern blot, đánh giá kiểu hình cây chuyển gen, định tính, định lượng chất mục tiêu.
3.11.2.3. Kiểm tra cây chuyển gen bằng Southern blot
Phân tích Southern blot gen cbfd2
Thông qua phân tích bằng PCR đã xác định được các cây đậu tương chuyển gen. Tuy nhiên để kiểm tra sự hòa nhập bền vững của các gen này vào bộ gen của cây, đồng thời xác định dòng chuyển gen riêng biệt, phân tích Southern blot, dựa trên lai phân tử với mẫu dò là đoạn trình tự của gen cbfd2 có kích thước 500 bp đã được hiện. DNA tổng số của các dòng chuyển gen, đối chứng âm (cây không chuyển gen), đối chứng dương (plasmid pITB-AST) được cắt bởi enzyme HindIII.
Kết quả trên hình 3.26 cho thấy ở đối chứng dương có hiện băng lai, đối chứng âm không hiện băng lai. Ở vị trí các dòng chuyển gen đều hiện diện băng lai với các kích thước khác nhau. Điều này chứng tỏ gen biến nạp đã được gắn chèn vào bộ gen cây ở những vị trí khác nhau, do đó các dòng này là các dòng chuyển gen riêng biệt. Ngoài ra, ở dòng D3 chỉ hiện diện một băng lai, cho thấy dòng này chỉ có một bản sao của gen được gắn chèn vào bộ gen. Các dòng còn lại đều có sự hiện diện từ 2-3 băng lai, chứng tỏ các dòng này được gắn chèn nhiều bản sao của gen vào bộ gen.
Nghiên cứu chuyển gen thông qua vi khuẩn A. tumefaciens thường tạo ít bản sao của gen biến nạp trong bộ gen cây. Do đó ít ảnh hưởng đến các tính trạng vốn có của cây chuyển gen. Hơn nữa việc có ít bản sao, tốt nhất là chỉ một bản sao của gen, giúp dễ chọn lọc dòng đồng hợp tử. Dòng này sẽ có sự di truyền và biểu hiện
gen ổn định hơn qua nhiều thế hệ. Trong 5 dòng chuyển gen được phân tích, có 2 dòng có 1 bản sao (D3, D5), 2 dòng có 2 bản sao (D2, D8) và 1 dòng có 3 bản sao (D9). Như vậy, nhìn chung các cây chuyển gen nhận được có số lượng bản sao của gen chuyển trong bộ gen không quá nhiều. Tuy nhiên, để biết ảnh hưởng của việc gắn chèn các bản sao này đến kiểu hình, sự sinh trưởng, phát triển cũng như khả năng sinh sản, cần có các đánh giá ở giai đoạn sau.
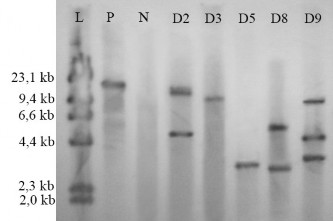
Hình 3.26. Kết quả phân tích DNA cây đậu tương chuyển gen bằng Southern blot
L. Thang chuẩn HindIII; P Đối chứng dương plasmid pITB-AST; N. Cây đậu tương không chuyển gen; D2, D3, D5, D8, D9 Các dòng cây đậu tương dương t nh PCR.
3.11.2.4. Kiểm tra tính kháng của cây chuyển gen với thuốc diệt cỏ Basta
Gen chọn lọc bar giúp chọn lọc được chồi, cây chuyển gen in vitro, đồng thời cũng giúp cây chuyển gen ex vitro kháng với thuốc diệt cỏ gốc glufosinate [112][130][178]. Do đó, thí nghiệm kiểm tra tính kháng của cây chuyển gen ngoài vườn ươm với thuốc diệt cỏ có thể làm cơ sở khẳng định sự hiện diện, biểu hiện của gen bar nói riêng cũng như góp phần dự đoán sự hiện diện, hoạt động của các cấu trúc gen đi kèm trong đoạn T-DNA. Dung dịch thuốc diệt cỏ Basta (100 mg/l glufosinate) được sử dụng để quét lên mặt trên lá của các cây chuyển gen (D2, D3, D5, D8, D9) và đối chứng cấy mô trồng ngoài vườn ươm. Kết quả được ghi nhận sau 7 ngày cho thấy lá của các cây chuyển gen hầu như không bị ảnh hưởng, vẫn giữ được màu xanh của diệp lục, trong khi đó lá của cây đối chứng bị mất màu xanh, cháy khô tại vùng quét Basta (hình 3.27). Điều này cho thấy gen kháng thuốc diệt cỏ bar gắn chèn trong bộ gen của các cây chuyển gen đã biểu hiện giúp cây kháng lại thuốc diệt cỏ ở nồng độ gây chết với cây đối chứng.
1 cm
a
b
Hình 3.27. Lá cây đối chứng và chuyển gen sau 7 ngày quét dung dịch Basta
a: lá cây đối chứng; b: lá cây chuyển gen
3.11.2.5. Kiểm tra sự di truyền của gen biến nạp
Trong 9 dòng đậu tương chuyển gen được kiểm tra bằng PCR trồng trong vườn ươm, chỉ 5 dòng có khả năng thích nghi, phát triển. Các dòng này cũng biểu hiện tính kháng với thuốc diệt cỏ Basta (100 mg/l glufosinate) và có sự hiện hiện các băng lai đặc hiệu trong thí nghiệm Southern blot với probe là đoạn gen cbfd2. Trong các dòng này, chỉ 2 dòng (D2, D8) có sự tạo hoa, trái, hình thành hạt (hình 3.28), 3 dòng còn lại (D3, D5, D9) không tạo trái, mà sau một thời gian phát triển lá vàng, rụng, cây chết dần. Một số nghiên cứu chuyển gen trên đậu tương cũng ghi nhận nhiều cây chuyển gen khi trồng trong vườn ươm không thể tạo hạt [130][181]. Nguyên nhân có thể do rễ những dòng chuyển gen này kém phát triển ảnh hưởng đến sự sinh trưởng, phát triển chung, ngoài ra vị trí gắn chèn của gen biến nạp cũng có thể gây ảnh hưởng đến khả năng sống của giao tử [181][182].
Sau khoảng 4 tháng phát triển trong vườn ươm, trái của 2 dòng D2 và D8 đến giai đoạn chín, vỏ trái khô, màu vàng. Dòng D2 thu được hai trái, mỗi trái 1 hạt, gồm 1 hạt màu đỏ (D2-1) và 1 hạt màu trắng (D2-2). Dòng D8 thu được 1 trái hai hạt, gồm 1 hạt đỏ (D8-1) và 1 hạt trắng (D8-2). Màu đỏ của các hạt thể hiện đậm và rõ ở phần rễ mầm, nhạt hơn ở phần phôi nhũ (hình 3.29). Nghiên cứu chuyển gen tạo astaxanthin trên nhiều cây khác nhau cho thấy sự tích lũy astaxanthin trên các bộ phận của cây chuyển gen dẫn đến màu của các bộ phận này chuyển sang đỏ như ở thuốc lá, táo, ngô, cà chua,… [7][31][60][62].
Bốn hạt của hai dòng D2 và D8 được gieo trồng trong vườn ươm để kiểm tra khả năng nảy mầm, sinh trưởng, phát triển của cây so với đối chứng. Trước tiên, các hạt chuyển gen và đối chứng được gieo nảy mầm trong đĩa petri có giấy thấm nước, ủ trong tối 4 ngày đến khi phần rễ mầm phát triển tốt. Sau đó, cây mầm được trồng trong chậu đất ở vườn ươm.