chủng Bt sinh tổng hợp 3 dạng độc tố chính là Cry, Cyt, Vip do các gene cry, cyt, vip mã hóa [21,25].
1.1.5.1.Gene cry
Gene cry mã hóa cho các độc tố Cry khác nhau. Đây là gene độc tố quan trọng nhất của Bt, tính đặc hiệu diệt côn trùng là do các protein độc do gene cry mã hóa. Các gene cry thường nằm trên các plasmid có hệ số copy thấp. Những nghiên cứu của Carlton và Gonzalez trên 21 loài phụ Bt đã cho thấy số lượng plasmid dao động từ 2- 12 trong các chủng nghiên cứu [11,20].
Theo công bố mới nhất trên website
http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/toxins2.html: đã phát hiện tới hơn 600 gene thuộc 70 họ gene cry của Bt.
Sau đây là các họ gene mã hóa cho protein tinh thể độc diệt một số bộ côn trùng gây hại phổ biến ở Việt Nam:
Bảng 1.1. Phân loại gene cry của Bt
Hình dạng tinh thể | Khối lượng phân tử protein (kDa) | Hoạt tính diệt côn trùng | |
cry1A- cry1L | Lưỡng tháp | 130 – 138 | Cánh vảy |
cry2A- cry2C | Cầu | 69 – 71 | Cánh vảy, hai cánh |
cry3A- cry3C | Lập phương | 73 – 74 | Cánh cứng |
cry4A- cry4D | Cầu | 73 – 74 | Hai cánh |
cry5 | Lưỡng tháp | 81 | Tuyến trùng |
cry8 | Lập phương | 130 | Cánh cứng |
Các gene cry còn lại | Đa dạng | 35 – 129 | Đa dạng |
Có thể bạn quan tâm!
-

 Nghiên cứu đặc điểm sinh học của một số chủng Bacillus Thuringiensis sinh Protein tinh thể diệt côn trùng cánh vảy - 1
Nghiên cứu đặc điểm sinh học của một số chủng Bacillus Thuringiensis sinh Protein tinh thể diệt côn trùng cánh vảy - 1 -
 Nghiên cứu đặc điểm sinh học của một số chủng Bacillus Thuringiensis sinh Protein tinh thể diệt côn trùng cánh vảy - 2
Nghiên cứu đặc điểm sinh học của một số chủng Bacillus Thuringiensis sinh Protein tinh thể diệt côn trùng cánh vảy - 2 -
 Gene Cry1C Và Dưới Loài Bacillus Thuringiensis Subsp . Aizawai
Gene Cry1C Và Dưới Loài Bacillus Thuringiensis Subsp . Aizawai -
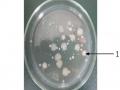 Phân Loại Các Chủng Bt Bằng Phản Ứng Huyết Thanh
Phân Loại Các Chủng Bt Bằng Phản Ứng Huyết Thanh -
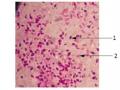 Nghiên cứu đặc điểm sinh học của một số chủng Bacillus Thuringiensis sinh Protein tinh thể diệt côn trùng cánh vảy - 6
Nghiên cứu đặc điểm sinh học của một số chủng Bacillus Thuringiensis sinh Protein tinh thể diệt côn trùng cánh vảy - 6 -
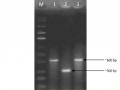 Nghiên cứu đặc điểm sinh học của một số chủng Bacillus Thuringiensis sinh Protein tinh thể diệt côn trùng cánh vảy - 7
Nghiên cứu đặc điểm sinh học của một số chủng Bacillus Thuringiensis sinh Protein tinh thể diệt côn trùng cánh vảy - 7
Xem toàn bộ 63 trang tài liệu này.
Gene cyt
Gene cyt mã hóa cho các độc tố Cyt cũng có bản chất protein và hình thành thể vùi như độc tố Cry, nhưng tính tương đồng amino acid giữa 2 độc tố là không đáng kể [34].
Gene cyt bao gồm 45 gene thuộc 3 lớp cyt1, cyt2 và cyt3 mã hóa protein gây độc với côn trùng và động vật không xương sống. Nhóm gene cyt mã hóa protein 27 kDa và chiếm khoảng 40-50% tinh thể, protein này có tác dụng gây độc với côn trùng bộ hai cánh và giun tròn.
Gene vip
Gene vip mã hóa cho các độc tố Vip (vegetative insecticidal protein). Độc tố Vip được Estruch và cộng sự phát hiện đầu tiên vào năm 1996, bao gồm Vip3A và Vip3B [8].
Độc tố Vip đã thu hút được sự quan tâm của rất nhiều nhà khoa học do khả năng diệt côn trùng phổ rộng, hoạt tính diệt sâu cao mà đặc biệt chưa phát hiện được côn trùng kháng độc tố này. Các nhà khoa học đã phát hiện được 85 gene vip thuộc 4 nhóm khác nhau, trong đó đáng chú ý là gene vip3A mã hóa protein 88 kDa có hoạt tính với nhiều côn trùng cánh vảy: Agortis ispilon, Spodoptera fugipera, Spodoptera exigua. Khi côn trùng ăn phải độc tố, Vip3A là nguyên nhân gây phân giải tế bào biểu mô ruột giữa.
1.1.6. Các loại độc tố của Bacillus thuringiensis
Bacillus thuringiensis có 4 loại độc tố: 3 ngoại độc tố và một nội độc tố: Ngoại độc tố ( - exotoxin), ngoại độc tố (-exotoxin), ngoại độc tố ( - exotoxin), nội độc tố (-endotoxin). Trong đó nội độc tố có tác dụng diệt côn trùng mạnh nhất.
Ngoại độc tố
Có khối lượng phân tử thấp, không bền nhiệt, có khả năng tan trong nước, tích tụ trong pha logarit của một vài loài phụ và được giải phóng ra môi trường khi tế bào tan rã. Ngoại độc tố có bản chất là một enzyme gọi là phospholipase hoặc leucithinase, có hoạt tính với một vài loài ong cắn lá đặc biệt là ong Tenthre dineda. Tác dụng độc của ngoại độc tố có liên quan tới sự phân huỷ phospholipid ở mô của côn trùng (nghiên cứu này được phát hiện bởi Toumanoff - năm 1953) [8, 11].
Ngoại độc tố α có tác dụng tiêu diệt Galleria mellonella. Ngoài ra còn có hiệu lực với các loài sâu thuộc bộ cánh vảy, cánh cứng.
Ngoại độc tố
Độc tố này được Halt và Arkawwa (1959) tìm ra khi nuôi ấu trùng ruồi nhà bằng thức ăn có chứa B. thuringiensis. Độc tố có khối lượng phân tử cao 707- 850 kDa, bền nhiệt, tan trong nước. Ngoại độc tố có tác dụng kìm hãm nuclease và RNA-polymease dẫn tới cản trở việc tổng hợp tRNA. Ngoại độc tố
có phổ hoạt tính rộng, kháng côn trùng thuộc bộ cánh vảy, côn trùng thuộc bộ cánh cứng và côn trùng thuộc bộ hai cánh [8,12].
Ngoại độc tố β rất có hiệu quả trong việc chống sâu non của côn trùng mẫn cảm, gây trì trệ trong việc chuyển hóa lột xác và có tác động đối với sâu trưởng thành phát triển từ các ấu trùng đã ăn phải độc tố dưới ngưỡng gây chết.
Qua nghiên cứu Federici và Wu cho thấy rằng chế phẩm Bt nếu chỉ sử dụng ngoại độc tố thì hiệu quả diệt côn trùng chỉ đạt 20% trong khi đó nếu bổ sung thêm nội độc tố thì hiệu quả diệt côn trùng sẽ tăng lên tới 75%.
Ngoại độc tố
Có khối lượng phân tử thấp, mẫn cảm với nhiệt độ, ánh sáng và không khí, có khả năng tan trong nước. Độc tố này thuộc nhóm phospholipase, có tác dụng lên phospholipid và giải phóng ra acid béo, rất mẫn cảm với nhiệt độ, ở nhiệt độ từ 600C trở lên trong vòng từ 10- 15 phút đã bị vô hoạt.
Nội độc tố
Có bản chất là một protein gồm 1180 gốc amino acid, các amino acid chủ yếu là glutamic và aspartic acid, chiếm trên 20% tổng số amino acid trong phân tử protein, đây cũng là một nguyên nhân gây ra điểm đẳng điện thấp. Lượng cysteine nhỏ hơn 20% tổng axit amin quy định sự không hoà tan của tinh thể, ngoài ra còn có các amino acid: arginine, threonine, leucine, isoleucine [14].
Ngoài protein tinh thể còn chứa các thành phần khác như: carbohydrate, tuy nhiên cũng có thể carbohydrate chỉ là thành phần phụ được pha tạp trong quá trình hình thành bào tử. Xét về thành phần hóa học thì nội độc tố chứa chủ yếu các nguyên tố C, H, O, N, S. Ngoài ra còn có Ca, Mg, Si, Fe và một lượng nhỏ Ni, Ti, Zn, Al, Cu, Mn. Nguyên tố P hầu như không có.
Nội độc tố : có đặc điểm không hòa tan trong các dung môi hữu cơ, chỉ tan trong môi trường có pH kiềm, không tan trong nước, rất bền nhiệt. Khi đun ở 650C trong một giờ không bị mất hoạt tính, chỉ khi đun ở 1000C trong 30-40 phút thì tinh thể bị mất tính độc. Mặc dù tinh thể độc có thể tan ở pH kiềm, nhưng không phải pH kiềm nào cũng thích hợp. Qua nghiên cứu các nhà khoa học nhận thấy rằng, pH quá cao hay quá thấp cũng đều ảnh hưởng tới độc tính của tinh thể. Tuy nhiên lúc mới tách ra khỏi bào tử, nó có thể hòa tan trong pH rất kiềm. Sự tổng hợp tinh thể độc và bào tử xảy ra gần như đồng thời trong khoảng 3 giờ sau pha cân bằng [8, 11, 16].
1.1.7. Cấu trúc của các nhóm độc tố tinh thể
Năm 1991, Li và cs đã công bố cấu trúc của nội độc tố của protein tinh thể diệt sâu Cry3A, từ đó đã mở rộng ra cho các độc tố khác. Các protein tinh thể có cấu trúc chung gồm 3 vùng riêng biệt:
Vùng 1: là một chuỗi polypeptide gồm 290 amino acid, đây là vùng đầu
N. Vùng này gồm 7 chuỗi xoắn , 6 trong 7 chuỗi này được sắp xếp thành hình 6 cạnh bao bọc xung quanh chuỗi xoắn thứ 5 kị nước. Mặt bên của chuỗi xoắn thứ nhất và thứ 7 là các amino acid kị nước và nó nằm liền kề với vùng 2.
Vùng 2: bắt đầu từ amino acid 291 đến amino acid 500. Vùng này gồm 3 tấm có cấu trúc tương đồng, song song và xếp ngược chiều nhau, trong đó tấm 1 và tấm 2 có tính tương đồng cao hơn. Mỗi tấm bao gồm 4 mảnh không song song, có cấu trúc chung. Hai mảnh ở phía trong tạo thành một dải kéo dài ra tận phía 2 mảnh ở phía ngoài. Tấm 3 có 3 mảnh và 1 chuỗi xoắn có chức
năng duy trì cấu trúc cơ bản của tấm 1 và 2. Vùng 2 là vùng không bền vững của độc tố [20].
Vùng 3: chứa những tấm dạng bánh kẹp Sandwich. Vùng này chứa đầu nhóm carboxyl và có cấu trúc bền vững [8].
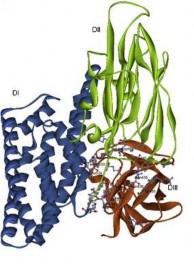
Hình 1.3. Mô hình cấu trúc không gian 3 chiều của protein độc tố Cry1Ac
1.1.8. Cơ chế tác động của protein tinh thể lên côn trùng
Cơ chế hoạt động của protein tinh thể của B. thuringiensis liên quan tới sự hòa tan tinh thể trong ruột giữa của côn trùng. Khi sâu ăn phải protein tinh thể của Bacillus thurigiensis, dưới tác động của men protease có tính kiềm (pH>10) trong ruột sâu, tinh thể độc tố bị hòa tan, và một nhân độc tố có khối lượng phân tử 60–65 kDa được hình thành và hoạt hóa. Độc tố đã hoạt hóa bám vào các phân tử cảm thụ đặc biệt nằm trên màng vi thể của tế bào thành ruột sâu. Sự gắn kết này ở ruột sâu làm thay đổi gradient điện hóa tạo thành các lỗ rò (kênh ion). Do đó phá hủy cân bằng áp suất thẩm thấu của màng tế bào, làm cho tế bào phồng lên, bị phân hủy dẫn đến sâu chết [8, 26].
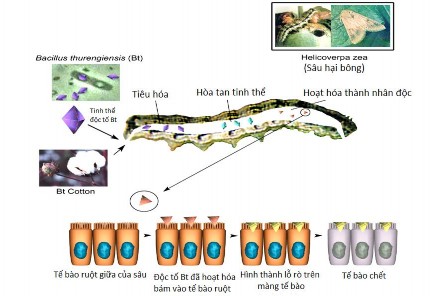
Hình 1.4. Cơ chế diệt sâu của vi khuẩn Bacillus thuringiensis [21]
1.1.9. Nghiên cứu và ứng dụng Bt ở Việt Nam
Ở Việt Nam thuốc trừ sâu sinh học Bt được ứng dụng đầu tiên tại Viện Bảo vệ Thực vật năm 1971. Nguyễn Văn Cảm và CS đã khảo nghiệm 5 loại thuốc trừ sâu sinh học Bt nhập nội từ Liên Xô vàTrung Quốc đã cho kết quả rất khả quan. Tuy nhiên, những nghiên cứu, sản xuất và ứng dụng Bt đầu tiên được Nguyễn Công Bình và CS thực hiện lần đầu tiên vào năm 1973 tại Viện Sinh vật nay là Viện Công nghệ sinh học. Cho đến nay Bt đã được nghiên cứu rộng rãi ở nhiều cơ sở tại Việt Nam như Viện Công nghiệp Thực phẩm, Viện Bảo vệ Thực vật, Viện Công nghệ sinh học [13].
Trên cơ sở phát hiện, tách dòng và đọc trình tự gene có trong các chủng Bt phân lập tại Việt Nam. Ngô Đình Bính và cs đã tách dòng gene và biểu hiện gene mã hóa tổng hợp protein Cry1C, Cry1D diệt sâu khoang từ các chủng Bt subsp. aizaiwai phân lập từ các mẫu đất của Hà Nội và Hà Tĩnh. Protein tái tổ hợp có hoạt tính diệt sâu cao hơn so với các chủng đối chứng [12].
Ở Việt Nam, những nghiên cứu về lĩnh vực chuyển gene kháng côn trùng vào cây trồng để tạo ra cây có khả năng kháng sâu bệnh mới đã được bắt đầu từ thế kỷ 20. Trong đó, đã có rất nhiều nghiên cứu chuyển gene cry1A kháng sâu vào cây trồng thông qua vi khuẩn Agrobacterium tumefaciens để tạo ra các
giống cây trồng mới có khả năng kháng sâu như cây đậu xanh, cây bông. Năm 2003, Phan Đình Pháp và CS đã chuyển gene cry1B vào cây lúa thông qua phương pháp sử dụng súng bắn gene, đến năm 2005, gene kháng sâu trên được chuyển vào cây cà tím thông qua Agrobacterium [12].
Những nghiên cứu trên đã tạo tiền đề cho các nghiên cứu tiếp theo trong việc ứng dụng các gene quý của Bt để tạo ra các cây trồng mới kháng được sâu bệnh cũng như sản xuất thuốc trừ sâu sinh học Bt.
1.1.10. Các yếu tố ảnh hưởng tới quá trình hình thành bào tử và tinh thể độc
Nghiên cứu các yếu tố ảnh hưởng tới sự hình thành bào tử và tinh thể độc, đóng một vai trò vô cùng quan trọng và có tác động mạnh tới hiệu quả của quá trình lên men sản xuất chế phẩm Bt. Nhiều nhà nghiên cứu đã nhận thấy rằng các yếu tố ảnh hưởng tới quá trình sinh trưởng phát triển của Bt đều ảnh hưởng tới quá trình sinh tổng hợp độc tố.
Nhiệt độ: Bt có khả năng sinh trưởng, phát triển trong khoảng nhiệt độ 20- 400C. Tuy nhiên nhiệt độ tối ưu cho sự sinh trưởng Bt là 28- 320C. Nếu nhiệt độ dưới 200C và trên 320C đều có ảnh hưởng lên quá trình hình thành bào tử. Nếu nuôi cấy ở 200C thì chu kì phát triển cần 64 giờ, trong khi đó nuôi cấy ở 350C thì thời gian cần cho chu kì phát triển giảm xuống còn 27 giờ và số bào tử tăng lên. Khi nuôi cấy Bt trên môi trường thạch ở nhiệt độ trên 400C thì Bt vẫn có khả năng phát triển nhưng bào tử lại không được hình thành. Thêm vào đó nhiệt độ cũng có ảnh hưởng lớn tới quá trình hình thành tinh thể độc, ở nhiệt độ dưới 140C sau 360 giờ nuôi cấy không quá 10% tinh thể được hình thành, nhưng ở nhiệt độ 160C sau 168 giờ nuôi cấy lượng tinh thể tăng lên đạt tới 15% [8].
pH: pH là một trong số các yếu tố có ảnh hưởng tới quá trình sinh trưởng Bt, ảnh hưởng đến hoạt tính của enzym, ảnh hưởng đến điện tích màng tế bào và sự hấp thu chất dinh dưỡng, ảnh hưởng tới khả năng cho nhận chất dinh dưỡng trong môi trường và tính độc của các vật chất có hại. Do đó pH quá cao hoặc quá thấp đều có hại tới Bt. Với pH dao động trong khoảng 5,8- 8,5 không
gây ảnh hưởng tới sự hình thành tinh thể độc. Tuy nhiên sự thay đổi lớn yếu tố pH lại làm thay đổi số lượng bào tử. Thường thì Bt phát triển tối ưu ở pH=7 [9].
Oxy: Bt là vi khuẩn hô hấp hiếu khí cho nên oxy là yếu tố không thể thiếu trong quá trình sinh trưởng Bt. Tùy từng giai đoạn sinh trưởng mà hàm lượng oxy cần cung cấp khác nhau. Ở các giai đoạn đầu của quá trình sinh trưởng, Bt cần nhiều không khí, do vậy không khí được cấp càng nhiều thì lượng sinh khối càng tăng và số lượng bào tử sẽ tăng. Tuy nhiên, ở các giai đoạn sau (giai đoạn logarit cho đến trở về sau) nếu cứ tiếp tục tăng hàm lượng không khí thì nó sẽ tác động lên quá trình sống của tế bào làm tế bào sớm tự phân giải, dẫn tới số lượng tinh thể độc giảm [8].
Các nguồn dinh dưỡng: Bulla và Arcas và cộng sự đã nghiên cứu và nhận thấy rằng các hợp chất amonium vô cơ như (NH4)2SO4 không có hiệu quả trong việc thúc đẩy quá trình sinh trưởng của Bt. Trong khi đó các nguồn nitơ hữu cơ như cao thịt, bột cá, bột đâụ tương, là yếu tố thúc đẩy quá trình sinh trưởng. Các chủng Bt khác nhau cần các amino acid khác nhau. Các nghiên cứu cũng chỉ ra rằng nguồn nitơ có ảnh hưởng rất lớn tới sinh khối Bt và các protein tinh thể độc. Ngoài ra các ion khoáng như Ca, Mg, K, cũng được coi là nguồn dinh dưỡng cần thiết cho sự sinh trưởng Bt.
Amino acid: một số amino acid như leucine, isoleucine có tác dụng ức chế sự sinh trưởng của Bt cũng như sự hình thành tinh thể độc, nhưng khi cho thêm axit amin valine vào thì sự ức chế bị mất đi. Ngoài ra, nếu bổ sung riêng rẽ threonine hoặc serine vào trong môi trường nuôi cấy Bt thì gây ức chế sự hình thành tinh thể độc nhưng nếu đưa cả hai loại đó vào trong môi trường thì sự ức chế bị mất. Tác dụng ức chế của amino acid serine cũng bị mất đi nếu như có mặt axit amin methionine. Acid – picoline và acid flucoacetate cũng ức chế việc tạo ra bào tử và sự hình thành tinh thể độc. Chất kháng sinh erythromycine ở nồng độ thấp không đủ để ức chế việc tạo ra bào tử và sự hình thành tinh thể độc [8].