Cordyceps là chi nấm nổi tiếng về sự đa dạng cũng như ứng dụng trong y học. Đã có 183 loài Cordyceps đã được ghi nhận [145] nhưng chỉ có 26 loài được tìm thấy trên các ký chủ Coleoptera, Diptera, Hemiptera, Heteroptera, Homoptera, Hymenoptera, Lepidoptera, và Orthoptera.Trong nghiên cứu này, Cordyceps spp. được tìm thấy trên 4 bộ ký chủ nhưng không tìm thấy trên Hemiptera (Bảng 3.1).
Isaria là chi nấm khá đa dạng với 72 loài được ghi nhận và đã được ứng dụng làm thuốc điều trị bệnh trong các bài thuốc cổ truyền ở một số vùng ở Nam Á [218]. Ký chủ chính của các loài Isaria là Coleoptera, Hemiptera, và Lepidoptera. Vì thế chủng nấm Isaria được tìm thấy ở bộ côn trùng Blattodae trong nghiên cứu là một ghi nhận mới.
Hiện có 270 loài Ophiocordyceps đã được ghi nhận trên thế giới. Hầu hết các loài hiện nay được chuyển từ các loài thuộc chi Cordyceps và một số chi liên quan như Beauveria, Paecoelomyces [165]. Loài nổi tiếng nhất của chi này là O. sinensis. Ký chủ chung của loài là Lepidoptera và Coleoptera [195].
Ngoài ra có thể thấy thành phần nấm ký sinh côn trùng ở khu vực nghiên cứu tương đối đa dạng. Tuy nhiên có sự khác biệt giữa hai khu vực nghiên cứu. Ở Khu bảo tồn thiên nhiên Copia, 10 mẫu nấm ký sinh côn trùng thu được trong 02 đợt thu mẫu được phân loại vào 4/6 chi nghiên cứu. Trong khi ở Vườn quốc gia Xuân Sơn, 14 mẫu nấm thu được trong 01 đợt thu mẫu được phân loại vào 6/6 chi nghiên cứu. Tuy nhiên, số lượng mẫu còn tương đối nhỏ (24 mẫu) nên không thể đánh giá được đầy đủ về đa dạng nấm ký sinh côn trùng ở hai khu vực này. Cần phải có nghiên cứu tiếp theo như tiến hành thực địa thu thập thêm nhiều mẫu trong những khoảng thời gian khác nhau, vị trí khác nhau để đánh giá đa dạng sinh học loài, đa dạng theo mùa, đa dạng về yếu tố địa lý…
Như vậy, từ 24 mẫu ký chủ thuộc các bộ côn trùng Blattodea, Hemiptera, Coleoptera, Hymenoptera, Lepidoptera thu thập được ở Khu bảo tồn thiên nhiên Copia và Vườn Quốc gia Xuân Sơn, chúng tôi đã tiến hành phân lập được 24 chủng nấm. Các chủng nấm đã phân lập được đa dạng phong phú
được phân loại vào 6 chi bao gồm chi Aschersonia, Purpureocillium, Beauveria, Cordyceps, Isaria, và Ophiocordyceps. Các nghiên cứu về tính đa dạng và các ứng dụng tiềm năng của các nhóm nấm này đã thu hút sự chú ý của các nhà khoa học trong nhiều thập kỷ [193, 206]. Tuy nhiên, các nghiên cứu về tính đa dạng COD và mối liên hệ giữa đa dạng loài và khả năng sản sinh COD của các loại nấm này còn khiêm tốn [141, 196].
3.2. Khả năng sinh tổng hợp cyclooligomer depsipeptide của các chủng nấm ký sinh côn trùng đã phân lập
Độc tố nấm nói chung và các hoạt chất COD nói riêng thường tìm thấy ở trong hệ sợi của nấm. Đối với các loại nấm ký sinh côn trùng hầu như các nghiên cứu chủ yếu tập trung vào nuôi cấy trên môi trường rắn và tách chiết các hoạt chất từ bào tử, hệ sợi hoặc thể quả tươi của nấm sau khi nuôi cấy. Tuy nhiên hiện nay có một số nghiên cứu về sản xuất các COD từ nấm bằng phương pháp nuôi cấy trong môi trường lỏng. Phương pháp này cho thấy sự ưu việt hơn so với lên men rắn về mặt thời gian nuôi cấy cũng như hàm lượng tích lũy COD trong sinh khối tế bào [168], [133]. Theo Supothina và cs khi sản xuất COD là beauvericin từ chủng nấm Isaria tenuipes BCC 31640 cho kết quả hàm lượng beauvericin thu được bằng cách lên men trong môi trường lỏng đạt gấp hơn 15 lần so với hàm lượng beauvericin trong thể quả khi nuôi cấy trong môi trường rắn [168]. Chính vì vậy, chúng tôi lựa chọn môi trường lên men lỏng để nghiên cứu khả năng sinh tổng hợp COD của các chủng nấm ký sinh côn trùng phân lập được. Khảo sát sự sinh tổng hợp COD của các chủng nấm đã phân lập bằng kỹ thuật sắc ký lỏng (Hình 3.26). Kết quả được thể hiện ở bảng 3.2.
Bảng 3.2. Khả năng sinh tổng hợp COD của các chủng nấm ký sinh côn trùng phân lập được ở khu vực nghiên cứu
Chủng | CDW (g/l) | COD (mg/g) | COD (mg/l) | |
1 | Aschersonia sp. XS01 | 3,97a±0,053 | 0,00a±0,000 | 0,00a±0,000 |
2 | Beauveria sp. CPA5 | 7,11c±0,123 | 1,09f±0,045 | 7,72f±0,131 |
3 | Beauveria sp. CPA15 | 7,23c±0,082 | 1,07f±0,074 | 7,7lf ±0,438 |
4 | Beauveria sp. CPA16 | 5,81b±0,050 | 4,98o±0,025 | 28,97i±0,275 |
5 | Beauveria sp. CPA44 | 11,05l±0,123 | 0,11a±0,012 | 1,18ab±0,089 |
6 | Beauveria sp. XS36 | 8,04e±0,070 | 0,58e±0,021 | 4,64de+±0,144 |
7 | Beauveria sp. XS37 | 11,55mn±0,041 | 0,51de±0,019 | 5,85ef±0,14 |
8 | Beauveria sp. XS38 | 10,23k±0,106 | 1,25f±0,033 | 12,8g+±0,329 |
9 | Beauveria sp. XS83 | 13,29p±0,057 | 2,77h±0,110 | 30,15i±0,945 |
10 | Cordycep sp. CPA3 | 6,08b±0,065 | 0,2ab±0,041 | 1,2ab±0,186 |
11 | Cordyceps sp. CPA13V | 11,77mn±0,180 | 2,61i±0,029 | 30,69ik±0,568 |
12 | Cordyceps sp. CPA14V | 9,84i±0,094 | 4,81n±0,074 | 47,29m±0,144 |
13 | Cordyceps sp. CPA31 | 14,07q±0,054 | 0,00a±0,000 | 0,00a±0,000 |
14 | Cordyceps sp. XS57 | 8,6f±0,102 | 0,46cde±0,009 | 3,98cde±0,041 |
15 | Cordyceps sp. XS67 | 8,48f±0,204 | 2,86k±0,110 | 24,27h±1,071 |
16 | Isaria sp. CPA40 | 10,42k±0,070 | 3,88m±0,021 | 40,39l±0,054 |
17 | Isaria sp. XS07 | 12,76o±0,029 | 1,88g±0,082 | 23,95h±0,775 |
18 | Isaria sp. XS66 | 8,62f±0,102 | 0,00a±0,000 | 0,00a±0,000 |
19 | Isaria sp. XS69 | 9,38h±0,140 | 3,48l±0,213 | 32,59k±1,102 |
20 | Isaria sp. XS71 | 11,85n±0,017 | 0,00a±0,000 | 0,00a±0,000 |
21 | Ophiocordyceps sp. CPA1 | 7,61d±0,090 | 0,62e±0,029 | 4,72de±0,122 |
22 | Ophiocordyceps sp. XS12 | 8,08e±0,143 | 0,06a±0,005 | 0,46a±0,029 |
23 | Ophiocordyceps sp. XS65 | 11,48m±0,016 | 0,00a±0,000 | 0,00a±0,000 |
24 | Purpureocillium sp. XS77 | 9,05g±0,176 | 0,45cde±0,029 | 4,1cde±0,127 |
Có thể bạn quan tâm!
-

 Nghiên cứu đa dạng và sinh tổng hợp Cyclooligomer depsipeptide của nấm ký sinh côn trùng tại Khu Bảo tồn thiên nhiên Copia và Vườn quốc gia Xuân Sơn - 10
Nghiên cứu đa dạng và sinh tổng hợp Cyclooligomer depsipeptide của nấm ký sinh côn trùng tại Khu Bảo tồn thiên nhiên Copia và Vườn quốc gia Xuân Sơn - 10 -
 Nghiên cứu đa dạng và sinh tổng hợp Cyclooligomer depsipeptide của nấm ký sinh côn trùng tại Khu Bảo tồn thiên nhiên Copia và Vườn quốc gia Xuân Sơn - 11
Nghiên cứu đa dạng và sinh tổng hợp Cyclooligomer depsipeptide của nấm ký sinh côn trùng tại Khu Bảo tồn thiên nhiên Copia và Vườn quốc gia Xuân Sơn - 11 -
 Định Loại Phân Tử Các Chủng Nấm Ký Sinh Côn Trùng Đ Phân Lập Ở Khu Vực Nghiên Cứu
Định Loại Phân Tử Các Chủng Nấm Ký Sinh Côn Trùng Đ Phân Lập Ở Khu Vực Nghiên Cứu -
 Kết Quả Xây Dựng Cây Phát Sinh Loài Dựa Trên Vùng Gen Its1-5.8S Rdna-Its2
Kết Quả Xây Dựng Cây Phát Sinh Loài Dựa Trên Vùng Gen Its1-5.8S Rdna-Its2 -
 Ảnh Hưởng Của Độ Ph Môi Trường Đến Sự Sinh Trưởng Và Khả Năng Sinh Tổng Hợp Cyclooligomer Depsipeptide Của Chủng C. Cateniannulata Cpa14V
Ảnh Hưởng Của Độ Ph Môi Trường Đến Sự Sinh Trưởng Và Khả Năng Sinh Tổng Hợp Cyclooligomer Depsipeptide Của Chủng C. Cateniannulata Cpa14V -
 Một Số Yếu Tố Ảnh Hưởng Đến Thu Nhận Cyclooligomer Depsipeptide
Một Số Yếu Tố Ảnh Hưởng Đến Thu Nhận Cyclooligomer Depsipeptide
Xem toàn bộ 228 trang tài liệu này.
Kết quả cho thấy, 19/24 (79%) chủng nấm phân lập được có khả năng sinh tổng hợp COD trung bình từ 0,11 - 4,98 mg/g. Hầu hết các chủng thuộc chi Isaria, Beauveria và Cordyceps đều sinh COD. Phát hiện này đã chứng minh rằng các loài nấm thuộc họ Cordycipitaceae rất có tiềm năng trong sản xuất COD và rất đáng để nghiên cứu [103, 168]. Trong nghiên cứu, Ophiocordyceps sp. CPA1, Ophiocordyceps sp. XS12 có khả năng sinh tổng hợp COD nhưng không tìm thấy COD ở chủng Ophiocordyceps sp. XS65. Tuy nhiên khả năng sinh tổng hợp COD cũng đã được ghi nhận ở một số loài nấm thuộc chi Ophiocordyceps như O. communis [65]. Có rất ít dữ liệu về khả năng sinh tổng hợp COD của chi Ophiocordyceps, các nghiên cứu hiện có chủ yếu tập trung vào ứng dụng trong y học của các loài nấm thuộc chi này [18]. Trong nghiên cứu này, tất cả các chủng thuộc chi Beauveria đều có khả năng sinh COD, trong khi 83,3% các chủng Cordyceps và 60% các chủng Isaria có khả năng này. Đây là kết quả rất đáng chú ý so sánh khả năng sinh tổng hợp COD của ba chi nấm Isaria, Beauveria và Cordyceps.
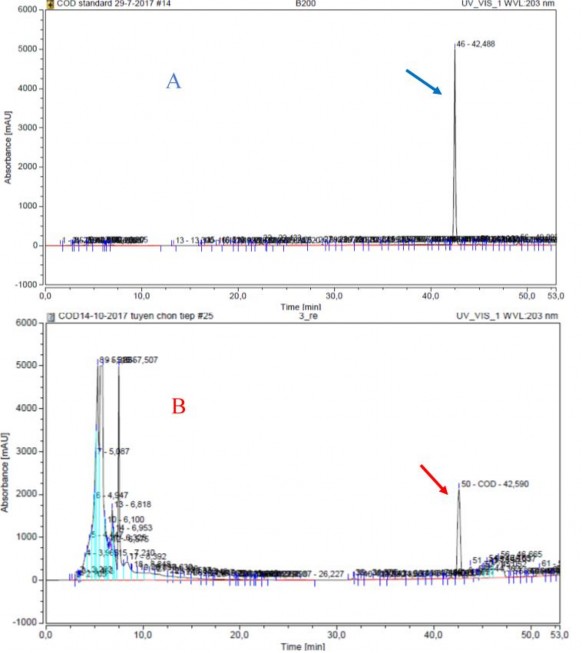
Hình 3.26. Sắc ký đồ HPLC UV 203 nm của chất chuẩn (A) và mẫu phân lập (B)
Bốn chủng Isaria sp. XS69, Isaria sp. CPA40, Cordyceps sp. CPA14V và Beauveria sp. CPA16 có khả năng sinh COD cao nhất, đạt 3,48, 3,88, 4,81, và 4,98 mg/g, trong khi sinh khối CDW tích lũy lần lượt đạt 9,38, 10,42, 9,84, và 5,81 g/l. Trong các chủng này thì chủng Cordyceps sp. CPA14V được tìm thấy trên ký chủ thuộc bộ côn trùng Blattodea có khả năng sinh 47.29 mg/l
sau 6 ngày nuôi cấy. Khả năng sinh tổng hợp tương đối cao so với kết quả trong các nghiên cứu của tác giả Logrieco (3,2 mg/g) [100], Luangsa-ard (36,9mg/l) [103]. Chủng nấm Cordyceps sp. CPA14V có tiềm năng, cần phải có thêm các nghiên cứu về công nghệ nuôi cấy và sản xuất COD từ chủng này. Các phát hiện của nghiên cứu này cho thấy rằng nấm ký sinh côn trùng khá đa dạng trong phân bố ký chủ và cũng phổ biến trong việc sinh tổng hợp COD [100, 183].
Như vậy, từ 24 mẫu nấm ký sinh trên 5 bộ ký chủ côn trùng được thu thập được ở Khu bảo tồn thiên nhiên Copia (Sơn La) và Vườn quốc gia Xuân Sơn (Phú Thọ), chúng tôi đã phân lập được 24 chủng nấm ký sinh côn trùng. Các chủng nấm phân lập được thuộc 6 chi gồm Aschersonia, Beauveria, Cordyceps, Isaria, Purpureocillium và Ophiocordyceps và có 19/24 chủng có khả sinh tổng hợp COD. Trong số đó hàm lượng COD cao nhất là của chủng Cordyceps sp. CPA14V (4,81 mg/g CDW hoặc 47,29 mg/l môi trường nuôi cấy trong 6 ngày). Chính vì vậy chúng tôi lựa chọn chủng nấm Cordyceps sp. CPA14V cho các nghiên cứu sâu hơn về đặc điểm hình thái, đặc điểm sinh học phân tử, điều kiện sinh tổng hợp, quy trình thu hồi và tinh sạch, xác định cấu trúc hóa học cũng như là hoạt tính sinh học của COD.
3.3. Định loại chủng nấm CPA14V bằng đặc điểm hình thái và sinh học phân tử
3.3.1. Đặc điểm hình thái của chủng nấm CPA14V
Chủng nấm Cordyceps sp. CPA14V được phân lập từ mẫu nấm ký sinh trên bộ Blattodea thu được tại Khu bảo tồn thiên nhiên Copia - Sơn La (Hình 1a).
Trạng thái vô tính ngoài tự nhiên: Bó sợi mọc thẳng, đơn độc, màu trắng kích thước 2,5-3 cm, phần đỉnh có phân nhánh kích thước 0,5-0,7 cm, có lớp bào tử dạng bột mịn, màu trắng. Các bào tử không có màng bao bọc, hình oval, kích thước 2,5-3 μm, bào tử không bắt màu với thuốc nhuộm nhuộm Melzer‟s (Hình 3.27a, 3.27b).
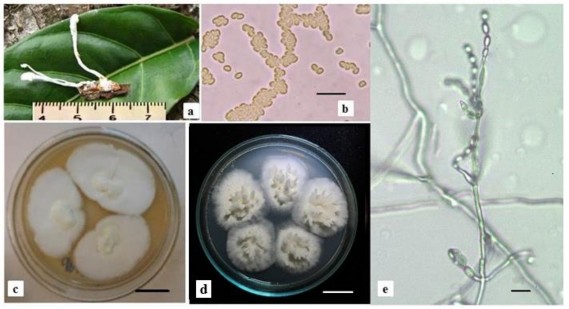
Hình 3.27. Đặc điểm hình thái của chủng Cordyceps sp. CPA14V
a) Trạng thái vô tính ngoài tự nhiên của chủng nấm Cordyceps sp. CPA14V; b) Bào tử chủng Cordyceps sp. CPA14V; c) Khuẩn lạc chủng Cordyceps sp. CPA14V trên môi trường Sabouraud; d) Khuẩn lạc chủng Cordyceps sp. CPA14V trên môi trường Czapek-Dox; e) Bào tử và cuống sinh bào tử chủng Cordyceps sp. CPA14V trên môi trường Czapek-Dox (scale bar: b,e=10 µm; c,d=2cm)
Đặc điểm nuôi cấy: Trên môi trường Sabouraud, khuẩn lạc nấm bông, dày, mịn, các sợi có màu trắng đục. Trên môi trường Czapek-Dox, khuẩn lạc có màu trắng, sau 20 ngày nuôi cấy ở điều kiện 25oC chiếu sáng 12 giờ hình thành bó sợi và bào tử vô tính (Hình 3.27b, 3.27d). Thể bình phình to thuôn dài có dạng cổ hẹp, kích thước 5-11,5 µm x 1,3-3,0 µm. Bào tử đính dạng chuỗi trên đó có các bào tử vô tính hình oval kích thước 2,5-3 µm (Hình 3.27e).
Đặc điểm hình thái trạng thái sinh sản vô tính của chủng nấm CPA14V đã phân lập giống với các đặc điểm của chi Paecilomyces, một trong những chi nấm ở trạng thái vô tính của các loài Cordyceps [85]. Trong phân loại nấm, các nhà khoa học vẫn sử dụng phương pháp phân loại truyền thống dựa
trên các đặc điểm hình thái quan sát được. Đặc biệt ở nấm ký sinh côn trùng, đặc điểm hình thái tế bào, đặc điểm nuôi cấy của các chủng nấm trong giai đoạn sinh sản vô tính là dữ liệu có giá trị trong định loại. Tuy nhiên, nếu chỉ dựa vào đặc điểm hình thái đôi khi chưa đủ để định loại chính xác một số loài nấm. Trong hơn 2 thập kỉ gần đây, nghiên cứu định loại nấm nói chung, nấm ký sinh côn trùng nói riêng cần nhiều dữ liệu phân tử, cùng với đặc điểm hình thái và hóa sinh để có kết quả đáng tin cậy hơn [41], [152]. Chính vì vậy, chúng tôi tiến hành định loại chủng nấm CPA14V bằng phương pháp sinh học phân tử.
3.3.2. Định loại chủng nấm CPA14V bằng phương pháp sinh học phân tử
Vật liệu di truyền của chủng nấm CPA14V đã tuyển chọn được tách chiết để khuếch đại trình tự gen ITS, LSU và Rpb1 bằng phản ứng PCR, sản phẩm được gửi sang công ty First BASE (Singapore) để tiến hành xác định trình tự nucleotide. Trình tự nucleotide của chủng nấm CPA14V được sử dụng để so sánh với các trình tự liên quan có sẵn trong Ngân hàng gen NCBI cho thấy rằng các trình tự tương tự nhất là các loài Cordyceps. Có 65 trình tự của Cordyceps, Isaria, Hypocrea, Samsoniella và Ophiocordyceps được chọn để phân tích (Bảng 2.2). Trình tự Ophiocordyceps được sử dụng làm trình tự gốc. Cây phát sinh chủng loại của chủng nấm CPA14V được xây dựng dựa trên trình tự các vùng gen ITS, LSU và Rpb1 bằng phương pháp Neighbor Joining với bootstrap 1000 sử dụng phần mềm Mega X, lần được các kết quả như hình 3.28, 3.29, 3.30.