Kháng sinh tác động trên màng nguyên tương
Kháng sinh có tác dụng gây rối loạn tính thấm của màng nguyên tương làm thoát các thành phần trong tế bào ra ngoài, hoặc tác động lên các phân tử cấu tạo màng nguyên tương, làm cho tế bào vi khuẩn không thể tiếp tục nhân lên được nữa.
Kháng sinh polimyxin, gramicidin, tyrocidin có cơ chế tác động này.
Kháng sinh ức chế quá trình sinh tổng hợp protein
Kháng sinh có tác dụng gây rối loạn sự tổng hợp protein do gắn với các ribosom ở các tiểu đơn vị 30 S hoặc 50 S dẫn đến cản trở sự kéo dài của chuỗi acid amin trong quá trình tổng hợp protein. Kháng sinh tác động theo cơ chế này có các Kháng sinh thuộc nhóm aminoglycosid, cloramphenicol, macrolid, lincosamid, streptogramin, Tetracyclin.
Kháng sinh ức chế sinh tổng hợp acid nucleic
Kháng sinh có tác dụng gây rối loạn sự tổng hợp ARN hoặc ADN của tế bào vi khuẩn như rifamycin, kháng sinh thuộc nhóm quinolon, novobiocin .
Kháng sinh ức chế quá trình sinh tổng hợp acid folic
Cơ chế tác động: acid folic là một coenzym đóng vai trò chủ đạo trong nhiều phản ứng chuyển hóa tạo acid amin, các purin và pyrimidin. Trong quá trình tổng hợp acid folic, tế bào vi khuẩn cần phải có acid para - aminobenzoic, acid glutamic và nhiều enzym trong đó có: dihydropteroat synthetase, dihydrofolat reductase. Một số thuốc kháng sinh đã tác động vào quá trình này làm cho các acid folic không được tạo ra, vì vậy nó đã tác động gián tiếp đến quá trình sinh tổng hợp các acid nucleic và protein của tế bào vi khuẩn như sulfamid, trimethoprim.
Có thể bạn quan tâm!
-

 Tỷ lệ nhiễm và mang gen kháng Cephalosporin thế hệ 3 và Quinolon của các chủng Klebsiella gây nhiễm khuẩn hô hấp phân lập tại Bệnh viện Nhi Trung ương, 2009 - 2010 - 1
Tỷ lệ nhiễm và mang gen kháng Cephalosporin thế hệ 3 và Quinolon của các chủng Klebsiella gây nhiễm khuẩn hô hấp phân lập tại Bệnh viện Nhi Trung ương, 2009 - 2010 - 1 -
 Tỷ lệ nhiễm và mang gen kháng Cephalosporin thế hệ 3 và Quinolon của các chủng Klebsiella gây nhiễm khuẩn hô hấp phân lập tại Bệnh viện Nhi Trung ương, 2009 - 2010 - 2
Tỷ lệ nhiễm và mang gen kháng Cephalosporin thế hệ 3 và Quinolon của các chủng Klebsiella gây nhiễm khuẩn hô hấp phân lập tại Bệnh viện Nhi Trung ương, 2009 - 2010 - 2 -
 Đề Kháng Thu Được Không Do Sinh Ra Enzym -Lactamase
Đề Kháng Thu Được Không Do Sinh Ra Enzym -Lactamase -
 Phát Hiện Các Đột Biến 158Delag, 5382Insc Và 6174Delt Bằng Real-Time Pcr (Scort Và Cs, 2006).
Phát Hiện Các Đột Biến 158Delag, 5382Insc Và 6174Delt Bằng Real-Time Pcr (Scort Và Cs, 2006). -
 Hình Ảnh Minh Hoạ Giải Trình Tự Một Đoạn Dna (Nguồn: Sanger Cequencing Read Display.gif)
Hình Ảnh Minh Hoạ Giải Trình Tự Một Đoạn Dna (Nguồn: Sanger Cequencing Read Display.gif)
Xem toàn bộ 134 trang tài liệu này.
1.4.2.Cơ chế đề kháng kháng sinh
1.4.2.1. Định nghĩa
Trong mối quan hệ giữa vi khuẩn và kháng sinh thì sự đề kháng được hiểu là khả năng chống đối của vi khuẩn đối với kháng sinh và hóa chất điều trị.
1.4.2.2. Các yếu tố làm cho vi khuẩn không chịu tác dụng của thuốc kháng sinh
Thuốc
- Liều lượng điều trị quá thấp hoặc thời gian điều trị quá ngắn
- Chọn sai thuốc: Do thầy thuốc chọn kháng sinh điều trị không phù hợp với ổ nhiễm trùng. Cho kháng sinh điều trị không dựa trên kết quả kháng sinh đồ.
- Thuốc hỏng: do mua phải thuốc hết hạn sử dụng, hoặc còn hạn nhưng bảo quản không tốt, hay chất lượng không bảo đảm nên hoạt tính kháng sinh không có.
- Kết quả kháng sinh đồ được xác định trên một chủng mà nó không phải là căn nguyên chính gây nhiễm trùng.
- Phối hợp hai kháng sinh có tính đối kháng nhau nên nó làm mất tác dụng của nhau.
Người bệnh
- Do cơ địa của người bệnh: trong một số trường hợp như giảm bạch cầu hạt, thiếu hụt globulin miễn dịch, suy giảm miễn dịch... Mặc dù kháng sinh được chọn là thích hợp và đến được ổ nhiễm khuẩn nhưng bệnh vẫn còn vì kháng sinh chỉ có tác dụng ức chế vi khuẩn nếu bệnh nhân có tình trạng miễn dịch kém sẽ không diệt được vi khuẩn.
- Tuổi tác của bệnh nhân: với những bệnh nhân còn quá non (trẻ sơ sinh) hoặc quá lớn tuổi do khả năng hấp thu thuốc kém và khả năng gắn thuốc vào protein huyết tương kém nên nồng độ kháng sinh ở trong máu và trong tổ chức rất thấp sẽ làm ảnh hưởng đến hiệu quả điều trị [9], [16].
1.4.2.3.Vi khuẩn đề kháng kháng sinh
Theo như định nghĩa trên thì sự đề kháng ở đây được hiểu là khả năng chống đối của vi khuẩn đối với kháng sinh và hóa chất điều trị. Khả năng đề kháng của vi khuẩn bao gồm khả năng tự nhiên (đề kháng tự nhiên) và khả năng thu được (đề kháng thu được) [70] .
Đề kháng tự nhiên
- Định nghĩa: đề kháng tự nhiên là khả năng đề kháng một số kháng sinh nhất định của một loài vi khuẩn nào đó mang đặc tính di truyền bình thường của loài.
- Ví dụ: Escherichia coli không chịu tác dụng của erythromycin, tụ cầu không chịu tác dụng của colistin và nalidixic acid, Pseudomonas aeruginosa bền vững với penicillin G.
Đề kháng thu được
- Định nghĩa: đề kháng thu được là khả năng đề kháng một hay nhiều kháng sinh do biến cố di truyền mà vi khuẩn từ chỗ không có trở thành có gen đề kháng.
- Những đề kháng thu được có thể là do:
. Đột biến trên NST: qua các thử nghiệm sao chép đĩa của Lederberg (1952) có thể giải thích được vi khuẩn đề kháng thuốc là do đột biến.
. Nhận được gen đề kháng qua các hình thức vận chuyển di truyền như: tiếp hợp khi hai vi khuẩn tiếp xúc trực tiếp với nhau, biến nạp khi vi khuẩn bị ly giải và giải phóng ra ADN tự do hoặc tải nạp nhờ phagiơ hoặc do một thành phần di truyền di động (transposons).
1.4.2.4. Cơ chế hóa sinh và di truyền học của đề kháng thu được ở vi khuẩn
Cơ chế sinh hóa của sự đề kháng thu được [18], [50]
- Làm giảm tính thấm của màng nguyên tương
- Làm thay đổi đích tác động
- Làm cho kháng sinh thoát ra ngoài màng tế bào vi khuẩn quá mức
- Tạo ra các enzym ức chế tác động của kháng sinh:
Cơ sở di truyền học của sự đề kháng thu được
Đề kháng thu được do đột biến trên NST
Đột biến thường xảy ra ở giai đoạn sao chép các bazơ purin và pyrimidin trong quá trình tổng hợp của ADN, các ADN mới được tổng hợp sẽ không còn giống với phiên bản của ADN thế hệ trước [48].
Đề kháng thu được do nhận được gen đề kháng qua các hình thức vận chuyển di truyền[12].
Biến nạp
Là hiện tượng một phần nhỏ ADN tự do của vi khuẩn cho vào được tế bào vi khuẩn nhận, gắn vào bộ gen của vi khuẩn nhận, quyết định tính chất mới của vi khuẩn này và tính chất này có thể di truyền.
Tải nạp
Tải nạp là hiện tượng chuyển gen từ vi khuẩn cho sang vi khuẩn nhận qua một phagiơ ôn hòa làm trung gian, phagiơ này gọi là phagiơ tải nạp.
Tiếp hợp
Tiếp hợp là hiện tượng chuyển gen từ vi khuẩn này sang vi khuẩn khác khi có sự tiếp xúc trực tiếp của vi khuẩn cho (được gọi là vi khuẩn đực) và vi khuẩn nhận (được gọi là vi khuẩn cái). Để có thể tiếp hợp được vi khuẩn cho phải có pili giới tính và toàn bộ hay một phần vật liệu di truyền sẽ được chuyển từ vi khuẩn cho sang vi khuẩn nhận qua cầu nối pili đó.
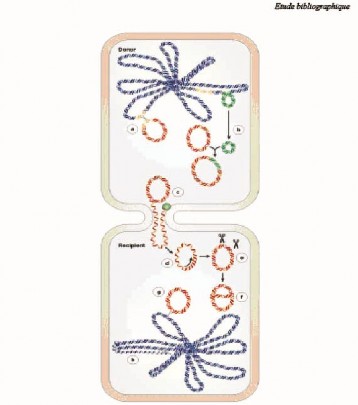
Hình 1.2. Sự lan truyền gen kháng thuốc bằng hình thức thông qua các R-plasmid [109]
Thành phần di truyền di động (transposons - gen nhảy)
Là một thành phần của một đơn vị sao chép hoặc là plasmid, nhiễm sắc thể hoặc là phagiơ nhờ hai chuỗi tận cùng ở hai đầu để chuyển vị trí từ một phân tử ADN này sang một phân tử ADN khác không cần có sự đồng dạng của hai phân tử ADN (nó có thể chuyển từ plasmid này sang plasmid khác, hay từ plasmid vào NST hoặc genom của một phagiơ tải nạp) [69].
- 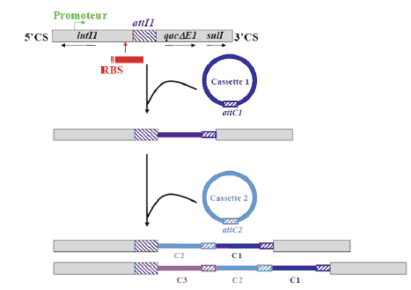
Hình 1.3. Đề kháng thu được do nhận được gene nằm trên Integron [109 ]
Cơ chế lan truyền gen đề kháng
Theo Wiedemann (1983) [121], gen đề kháng lan truyền trên bốn phương
diện:
- Trong tế bào (intracellular): qua các sự kiện tái tổ hợp kinh điển hay tái tổ
hợp bất hợp pháp tức là chuyển vị trí nhờ transposon.
- Giữa các tế bào (intercellular): qua các hình thức vận chuyển di truyền như: tiếp hợp, dẫn truyền, biến nạp, tải nạp. Tuy là hiện tượng ít gặp nhưng là tiền đề cho những hậu quả tiếp theo có ý nghĩa quan trọng về mặt số lượng.
- Trong quần thể vi khuẩn (microbiotop): thông qua sự chọn lọc và giữ lại những cá thể đề kháng trong vi hệ đường ruột hay trên da hoặc niêm mạc của bệnh nhân khi người này dùng kháng sinh.
- Trong quần thể đại sinh vật (macrobiotop): có thể thành dịch do lây lan từ người này sang người khác qua các dụng cụ thăm khám, dụng cụ phẫu thuật, thức ăn, không khí... bị nhiễm vi khuẩn đề kháng. Đây là điểm đặc biệt cần quan tâm ở các bệnh viện, nhằm phòng ngừa nhiễm khuẩn bệnh viện.
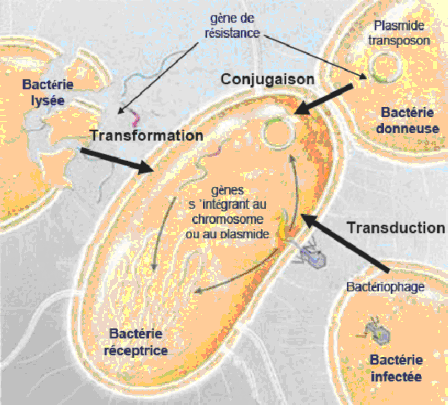
Hình 1.4. Giới thiệu các cơ chế khác nhau của sự lan truyền ngang các gene mã hóa sự kháng thuốc của vi khuẩn [109]
1.4.2.5. Các cơ chế đề kháng kháng sinh họ vi khuẩn đường ruột với kháng sinh nhóm beta - lactam
Đề kháng tự nhiên
Một số chủng thuộc họ vi khuẩn đường ruột có khả năng đề kháng tự nhiên với kháng sinh nhóm -lactam, nó mang đặc tính di truyền bình thường của loài. Nghiên cứu trên những chủng vi khuẩn họ đường ruột với các kháng sinh khác nhau trong nhóm -lactam đã cho phép người ta xác định được những kiểu cách đề kháng tự nhiên khác nhau của chúng [112].
Đề kháng tự nhiên do sinh ra enzym -lactamase
Đề kháng tự nhiên không do sinh ra enzym -lactamase
Một số vi khuẩn đường ruột đề kháng tự nhiên với kháng sinh thuộc nhóm - lactam là do thay đổi tính thấm của màng tế bào vi khuẩn đối với kháng sinh thuộc nhóm này [63], [82].
Đề kháng thu được
Các chủng vi khuẩn họ đường ruột có thể đề kháng với kháng sinh nhóm - lactam theo cơ chế đề kháng thu được (do biến cố di truyền mà vi khuẩn từ chỗ không có trở thành có gen đề kháng). Sự đề kháng này làm thay đổi các kiểu cách đề kháng tự nhiên của chúng. Những gen đề kháng thu được này có thể làm cho vi khuẩn sinh ra enzym -lactamaza, làm thay đổi tính thấm của màng tế bào hoặc làm thay đổi đích bám của kháng sinh lên màng tế bào vi khuẩn (làm thay đổi đích bám PBP).
Đề kháng thu được làm cho vi khuẩn sinh ra enzym -lactamase
Vi khuẩn sinh ra enzym penicillinase do nhận được gen đề kháng nằm trên plasmid
- Sinh ra penicillinase ở mức độ cao: những vi khuẩn họ đường ruột khi nhận được gen đề kháng nằm trên plasmid nó có thể sinh ra enzym penicillinase ở mức độ cao, những enzym này đã được xác định là các -lactamase phổ rộng ( TEM-1, TEM-2, SHV-1...) [72]. Những chủng này sẽ kháng lại các kháng sinh: penicillin, cephalosporin thế hệ I và II (trừ cephoxitin). Các kháng sinh cephalosporin thế hệ III, các monobactam và các carbapenem thì không bị phá hủy bởi enzym này. Các enzym này bị ức chế bởi các chất ức chế -lactamase.
- Sinh ra -lactamase phổ mở rộng(ESBL): một số vi khuẩn họ đường ruột
khi có khả năng sinh ra các -lactamase phổ rộng và nhờ đột biến nên đã mở rộng tác dụng của các enzym này tới kháng sinh cephalosporin thế hệ III (trừ các kháng sinh: cephamycin, cefoxitin, cefotetan, lactamoxef và carbapenem là không bị phá hủy bởi enzym này) như: enzym CTX-1/ TEM-3, TEM-4, CAZ-1/TEM-5, SHV-2,
SHV-3... [106], [95]. Các enzym -lactamaza phổ mở rộng này thường gặp ở loài Klebsiella pneumoniae và chúng bị ức chế bởi các chất ức chế -lactamase. Ở một số loài như E. coli, E. cloacae... sinh ra enzym này nhờ có gen mã hóa nằm trên plasmid [83], [86], [94].
- Sinh ra -lactamase kháng lại chất ức chế -lactamase: một số chủng đường ruột có những bước đột biến sinh ra các enzym mới đề kháng với chất ức chế
-lactamase, đã tìm thấy ở E. coli theo Vedel (1992) [111].
Những chủng đường ruột sinh ra enzym penicillinase do nhận được gen đề kháng nằm trên plasmid thường cũng kháng với các kháng sinh thuộc nhóm aminosid, chloramphenicol, trimethoprim và sulfamid do những gen mã hóa sự đề kháng với các nhóm kháng sinh này cùng nằm trên một plasmid [100].
Vi khuẩn tăng cường sản xuất ra enzym penicillinase do gen đề kháng nằm trên nhiễm sắc thể
Tăng cường sản xuất enzym penicillinaza ở K. oxytoca: những chủng của loài K. oxytoca kháng kháng sinh thuộc nhóm -lactam ở mức cao vì có sự tăng cường sản xuất enzym penicillinase nhờ gen mã hóa nằm trên NST theo tác giả Lambia và cộng sự năm 1986 [73].
Đề kháng thu được làm cho vi khuẩn sinh ra enzym cephalosporinase do gen mã hóa nằm trên nhiễm sắc thể
Sinh ra enzym cephalosporinase nhờ gen mã hóa nằm trên NST thường gặp ở loài E. cloacae, S. marcescens, E. coli trong họ vi khuẩn đường ruột:
- Tăng cường sản xuất enzym cephalosporinase: những loài này bình thường chúng kháng kháng sinh nhóm -lactam theo cơ chế đề kháng tự nhiên, do sinh ra enzym cephalosporinase cảm ứng và khi có sự đột biến ở gen điều hoà trong quá trình sinh tổng hợp enzym, nên đã tăng cường sản xuất enzym này. Cơ chế này được mô tả bởi hai tác giả Nicolas (1987) và Cole (1988) [37], [90].
- Tăng cường sản xuất enzym cephalosporinase ở loài E. coli là do hậu quả của sự khuếch đại gen hoặc do có sự thiết lập những gen điều hòa mới theo Normark (1977) và Cole (1988) [88], [37].