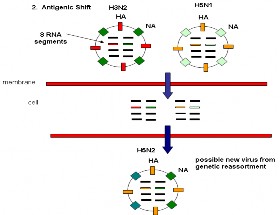
NA, hoặc cả hai gen của các chủng virus cúm A đã thích nghi người, để tạo ra một biến chủng virus mới thích ứng lây nhiễm dễ dàng người, gây ra nguy cơ của một đại dịch cúm mới và đặt ra một định hướng mới trong phòng chống (Hilleman, 2002; Guan và cs, 2002; Li và cs, 2004; Kim và cs, 2008).
TRỘN KHÁNG NGUYÊN 8 phân đoạn gen Màng tế bào TẾ BÀO Chủng virus mới được tạo ra do hòa trộn các phân đoạn gen | |
Hình 1.6. Sơ đồ minh họ đ t biến đi m củ c c phân đoạn gen i c A | Hình 1.7. Sơ đồ minh họa hiện tượng tr n kh ng ng yên củ i c A/H5N1 à H3N2 |
Có thể bạn quan tâm!
-
 Nghiên cứu một số đặc tính sinh học của vi rút cúm A/H5N1 Clade 7 phân lập ở Việt Nam - 2
Nghiên cứu một số đặc tính sinh học của vi rút cúm A/H5N1 Clade 7 phân lập ở Việt Nam - 2 -
 Bi Đồ Bi U Diễn Dịch C Gi Cầ Do I C A/h5N1 Theo Thời Gi N (Cục Th Y, 2012)
Bi Đồ Bi U Diễn Dịch C Gi Cầ Do I C A/h5N1 Theo Thời Gi N (Cục Th Y, 2012) -
 M Ph Ng Cấ T C Kh Ng Ng Yên H E L Tinin À Neuraminidase (Www.aht.org.uk)
M Ph Ng Cấ T C Kh Ng Ng Yên H E L Tinin À Neuraminidase (Www.aht.org.uk) -
 C C Genotype Củ I C Gi Cầ A/h5N1 Đ C Lực Cao (Li À C , 2007)
C C Genotype Củ I C Gi Cầ A/h5N1 Đ C Lực Cao (Li À C , 2007) -
 Kết Quả Tiê Phòng Vacxin C Gi Cầ Chương T Ình Q C Gia
Kết Quả Tiê Phòng Vacxin C Gi Cầ Chương T Ình Q C Gia -
 Bảng Tổng Hợp Liệ Tính To N Theo Phương Ph P Reed-Muench
Bảng Tổng Hợp Liệ Tính To N Theo Phương Ph P Reed-Muench
Xem toàn bộ 162 trang tài liệu này.
1.3.3. Đặc đi tiến hó à hình thành genotype củ i c gi cầ gi i đoạn 1996-2008
1.3.3.1. Sự tiến hó củ i c A/H5N1 à phân loại Clade dự t ên phân tích hệ phả gen HA.
Năm 1996, H N1 được phân lập từ ngỗng tại một ổ dịch Quảng Đông (Trung Quốc) và chủng này được coi là chủng nguyên thủy tạo nên các dòng virus gây bệnh cúm gia cầm trong 12 năm qua (Xu và cs, 1999). Chủng virus nguyên thủy này, lúc đó cung cấp nguồn gen HA(H ) cho tiến trình tái tổ hợp tạo nên các biến chủng gây dịch bệnh trên gia cầm và người Hong Kong năm 199 , và nguồn gen khung khác của virus cúm A/H5N1 Hong Kong được kiến tạo từ virus cúm A có chim cút (Guan và cs, 2002). iêng nguồn gen NA(N1) còn chưa biết được lấy từ đâu, nhưng cấu trúc của gen có hiện tượng xóa đi nucleotide mã hóa cho 19 amino acid, tại vùng đầu N của protein neuraminidase, và đột biến “xóa gen” này của N1 có liên quan đến tính thích ứng của virus cúm từ thuỷ cầm lên gia cầm trên cạn và người ( atrosovich và cs, 1999). Đối với gen HA(H ) đột biến giãn n chuỗi nối giữa HA1 và HA2 mã hóa cho các amino acid kiềm
(Arginine và Lysine) có liên quan đến tiến trình tăng cường độc lực, và các chủng thuộc dòng Quảng Đông (Guangdong-like sublineage), các amino acid thông thường là -RRRKK- (Claas và cs, 1998; atrosovich và cs, 1999).
Sau một năm gây bệnh tại Hong Kong, do toàn bộ đàn gia cầm bị tiêu diệt, virus cúm A/H5N1 nguyên thủy gốc Quảng Đông không còn gia cầm cạn để gây bệnh, người ta tư ng chúng đã biến mất, nhưng thực tế chủng nguyên thủy này vẫn tiếp t c tồn tại trong ngỗng vùng Nam Trung Quốc, tr thành nguồn gen tái tổ hợp hình thành biến chủng mới (Cauthen và cs, 2000; Wasilenko và cs, 2008). Trong các năm 199 - 2002, một số biến chủng virus cúm A/H5N1 mang nhiều đặc tính kháng nguyên khác nhau của subtype H được hình thành tạo nên nhóm kháng nguyên (clade) 1 có độc lực cao với gà nhưng thấp đối với vịt, để rồi bị đào thải trong những năm 2001 - 2002 (Guan và cs, 2002; Lee và cs, 200 ). Tiếp t c, trong năm 2002 - 2003, gen HA(H ) có những đột biến mới do hậu quả của hiện tượng lệch kháng nguyên (antigenic drift), để rồi tạo nên biến chủng có tính gây bệnh cực kỳ cao, đặc biệt đối với vịt, và lây sang người (Guan và cs, 2002; Sturm- amirez và cs, 200 ). Đặc tính thích ứng gây bệnh trên người càng ngày càng cao dần, cùng với độc lực tăng cường đối với đa vật chủ bao gồm vịt, gà, ngan, ngỗng, chim cút, chim hoang dã và người, để rồi hình thành nhiều biến chủng xâm nhập các nước phía Nam châu Á trong đó có Việt Nam, Thái Lan, Indonesia, Lào, Campuchia…(Guan và cs, 2002; Pantin-Jackwood, 2007).
Có thể nói, sau giai đoạn 199 - 2003, virus cúm A H N1 đã đạt đến mức độ hoàn thiện về đặc tính gây bệnh và thích ứng đa vật chủ, tr nên mối nguy cơ gây bệnh rất cao đối với gia cầm và người trong các năm 2004 - 200 (Lee và cs, 200 ; Smith và cs, 2006). Tuy nhiên, xét về di truyền học và tính kháng nguyên, các chủng H N1 giai đoạn 199 - 2002 vẫn mang tính đồng nhất kháng nguyên cùng với chủng nguyên thuỷ A Gs Gd 1 96 của Quảng Đông (Chen và cs, 2004), và bắt đầu phân hóa giai đoạn dịch cúm ác liệt xảy ra năm 2003 - 2005 (De Jong và Hien, 2006). Sự xuất hiện của genotype Z với tính gây bệnh ác liệt trong những năm này các nước Đông Nam Á là b ng chứng của sự đột biến “lệch kháng nguyên” của cúm A H N1 (Lee và cs, 200 ; WHO, 2004).
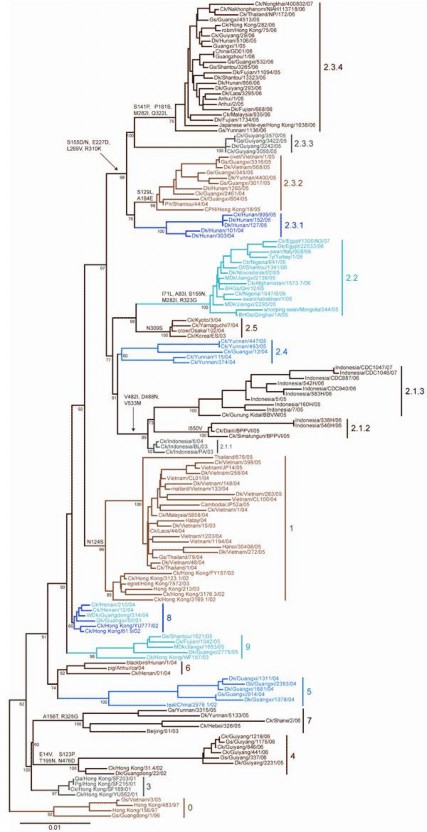
Hình 1.8. Cây phả hệ dự t ên gen HA c c i c A/H5N1 đ c lực cao (WHO, 2008)
Cuối năm 200 , có nhiều dưới dòng của virus cúm A/H5N1 cùng lúc được hình thành, đó là sự xuất hiện phân dòng Thanh Hải (Qinghai and Qinghai-like sublineage) và phân dòng Phúc Kiến (Fujian and Fujian-like sublineage), tràn ngập châu Á bao gồm Trung Quốc, Hong Kong, Việt Nam, Indonesia, Thái Lan (Lee và cs, 200 ; Sim và cs, 200 , Simmons và cs, 200 ), tràn sang Trung Á, châu Âu và châu Phi có tính gây bệnh cao đối với người (Ducatez và cs, 200 ; Pantin-Jackwood, 200 ; Zhao và cs, 2008). Các chủng thuộc dưới dòng Phúc Kiến có cấu trúc gen NA(N1) không thay đổi nhiều, nhưng gen HA(H ) có motif amino acid vùng nối của điểm cắt protease là -RRRK-, giảm mất một Lysine
(K) so với các chủng dưới dòng Quảng Đông (Simmons và cs, 200 ), và do vậy kể từ 2006 đến nay, nhiều chủng dòng virus cúm A/H5N1 cùng tồn tại gây bệnh, trong đó có nước ta (Dung Nguyen T và cs, 2008). Trong các năm 2006 - 2008, tuy bình diện dịch cúm gia cầm không ác liệt như những năm 2003-200 , nhưng do xuất hiện nhiều chủng A H N1 có biến động kháng nguyên và độc lực, vấn đề dịch tễ học cúm A/H5N1 có thể đã tr nên phức tạp hơn (Zhao và cs, 2008; Gambotto và cs, 2008).
Phân tích phát sinh loài được thực hiện b ng nhiều cách tiếp cận trên dựa trên tất cả các trình tự gen H5 HA (đã được công bố) tiến hóa từ virus H5N1 A/goose/Guangdong 96. Kết quả ban đầu cho thấy virus H N1 đang lưu hành có thể được nhóm lại thành nhiều loại "clades" virus dựa trên các đặc điểm phát sinh loài và trình tự tương đồng của gen HA(H5).
Đến nay, hệ thống thống nhất danh m c H N1 đã xác định được 10 clades ban đầu riêng biệt của virus (số 0-9) (Nhóm công tác tiến hóa H N1 WHO OIE
FAO, 2008), chúng được gọi là clades lớp thứ nhất. Việc xác định clade được cần đáp ứng ba tiêu chí định nghĩa nhánh c thể sau đây được phát triển b i nhóm công tác tiến hóa H5N1 WHO / OIE / FAO:
- N m trong cùng một nốt nhánh (clade);
- Tạo thành nhóm đơn ngành với giá trị bootstrap ≥ 60 tại nốt nhánh (dựa trên giá trị boostrap 1000 lần);
- Tỷ lệ phần trăm khoảng cách trung bình so sánh theo cặp nucleotide giữa các clade > 1, % và trong cùng clade <1, %.
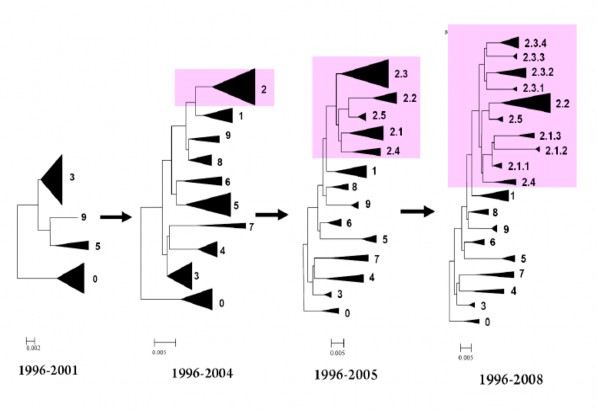
Hình 1.9. Sự tiến hó củ c c cl de i A/H5N1 theo thời gi n (WHO, 2010)
Trong giai đoạn 1996-2001, virus cúm gia cầm độc lực cao H N1 chủ yếu chỉ lưu hành phía Nam Trung Quốc. Giai đoạn này ghi nhận có 4 clade virus A H N1 khác nhau là clade 0, clade 3, clade và clade 9, trong đó clade 0 là virus thuỷ tổ A Goose Guangdong 96 và những virus khác cùng loại. Qua sơ đồ tiến hoá của virus cúm gia cầm H N1 độc lực cao (hình 1.9) qua các giai đoạn khác nhau, có thể nhận thấy sự biến đổi liên t c của virus cúm và đặc biệt là có những clade virus tiếp t c tiến hoá và phân loại thành các nhánh ph như clade 2: Năm 2004 chỉ có 1 lớp clade 2; Đến năm 200 clade 2 đã tiến hoá thành nhánh ph thuộc lớp thứ 2 từ clade 2.1 – 2.5, rồi đến năm 2008 chúng lại tiến hoá thành lớp thứ 3. Hiện nay các chuyên gia quốc tế đang tiếp t c tổng hợp và phân tích sự phát sinh loài của virus cúm và có thể sự phát sinh loài còn đa dạng hơn nữa.
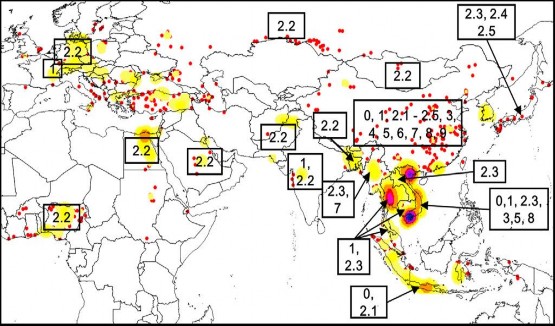
Hình 1.10. Sự phân b củ c c cl de i c gi cầ t ên thế giới từ 2003- 2009 ( feiffe à c , 2011 .
Cho đến nay tất cả 10 clade của virus cúm gia cầm độc lực cao H N1 đã được phát hiện và phân lập khu vực Đông Á có thể cùng hoặc các thời điểm khác nhau (Pfeiffer và cs, 2011). Về trình tự gen, hầu hết các virus cúm A/H5N1của khu vực Đông Á đều bắt nguồn từ Trung Quốc và Hong Kong và cũng được tìm thấy tại Nhật Bản và Hàn Quốc. ột số clade không phát hiện thấy Đông Á là clade 2.1.2 và 2.1.3. Dường như chúng đã tiến hóa Indonesia sau khi tổ tiên của chúng xâm nhập từ tỉnh Hồ Nam của Trung Quốc năm 2002- 2003 (Wang và cs, 2008). ột số clade cho đến nay chỉ mới phát hiện thấy Trung Quốc (ví d các clade 4, clade 6 và clade 9) trong khi các clade khác có liên quan đến các v dịch chính (clade 1 khu vực sông ê Kông từ năm 2004- 200 , clade 2.1 Indonesia năm 2004, clade 2.2 lan rộng từ châu Á sang châu Âu và châu Phi từ năm 200 , và clade 2.3.4 đã tấn công các nước Việt Nam, Lào và Thái Lan từ năm 200 ) đều đã được phân lập Trung Quốc trước khi được phát hiện thấy những nơi khác. Clade cũng mới chỉ xuất hiện Trung Quốc từ năm 200 tr về trước.
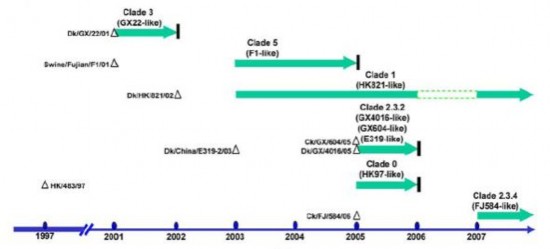
Hình 1.11. Thời gian xuất hiện củ c c cl de H5N1 ở Việt Nam từ 2001-2007 (Wallace à c . 2007).
Virus A H N1 gây nên dịch cúm gia cầm Việt Nam từ cuối năm 2003. Tuy nhiên, qua điều tra, người ta đã từng phát hiện thấy virus A/H5N1 chợ gia cầm sống Việt Nam từ năm 2001 (Nguyen và cs, 2001). Phân tích phát sinh loài các virus cúm đã từng phát hiện Việt Nam từ năm 2001-200 , đã thấy ít nhất có 6 clade HA khác nhau của virus cúm A/H5N1 độc lực cao từng xuất hiện và lưu hành Việt Nam (Wallace và cs. 200 ). Sáu clade HA này khi phân tích theo hệ thống danh pháp quốc tế rất tương đồng với các virus A/H5N1 tiền thân đã được xác định trước đó: clade 0 - giống virus HK97(A/HK/483/97); clade 1 - giống virus HK821(A/Dk/ HK/821/02); clade 2.3.2 - giống virus E319 (A/Dk/China/E319-2/03); clade 2.3. 4 - giống virus FJ584 (A/Ck/Fujian/584/05); clade 3-giống virus GX22 (A/Dk/GX/22/01); clade 5-giống virus F1(A/swine/Fujian/F1/01). Sáu clade virus này n m cùng nhóm với các virus tiền thân được phân lập trước đây tại Trung Quốc và Hồng Công và các virus tiền thân này có thể được coi là tổ tiên của các dòng virus cúm gia cầm độc lực cao xâm nhập vào Việt Nam. Thời điểm phân lập được các clade virus A/H5N1 từ gia cầm chỉ ra r ng một số dòng virus đã lưu hành trong vòng một năm hoặc ngắn hơn, trong khi những dòng khác tiếp t c lưu hành tại Việt Nam hơn lâu hơn. Ví d , sau khi được phát hiện lần đầu tiên trong năm 2003, virus clade 5 lưu hành tại Việt Nam khoảng 1 năm và được thay thế b ng clade 1 virus trong năm 2004 (Wallace và cs. 200 ).
Cho đến năm 200 , có 2 clade virus A H N1 chủ yếu lưu hành Việt Nam là clade 1 và clade 2.3.4. Clade 1 đã xuất hiện từ năm 2003 2004 phân bố trong cả nước, nhưng chủ yếu chỉ còn lưu hành phía miền Nam từ năm 200 cho đến nay; còn clade 2.3.4 xuất hiện từ năm 200 , trước đó đã lưu hành rộng rãi phía Nam Trung Quốc và xuất hiện phân bố và lưu hành chủ yếu miền Bắc Việt Nam, và đã thay thế cho clade 1 kể từ năm 200 (Dung Nguyen T và cs, 2008; Wallace và cs. 200 ), nhưng từ 2010 đến nay không còn phát hiện được nữa (C c Thú y)
Từ 2008-2010, dịch cúm gia cầm H N1 độc lực cao vẫn tiếp t c xảy ra và hầu hết các mẫu virus A H N1 đều được thu thập để phân tích và giám sát sự biến đổi của chúng. Kết quả cho thấy trong giai đoạn này, các virus A H N1 Việt Nam, đặc biệt là phía Bắc chủ yếu thuộc về clade 2.3.4, đồng thời các virus trong clade này tạo thành 1 lớp thứ tư thành 4 nhóm (2.3.4.1- 2.3.4.4)(Nguyen, 2010).
Từ 2009-2010, virus A/H5N1 thuộc clade 2.3.2 cũng đã xuất hiện Việt Nam và về mặt di truyền những virus này tương tự các virus A H N1 chim hoang và gia cầm của nhiều nước Đông Âu, Hồng Công, Trung Quốc, Hàn Quốc.
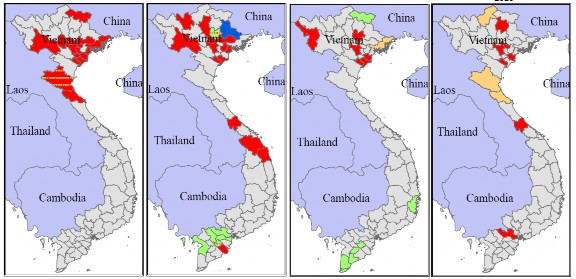
2008 | 2009 | 2010 |
Hình 1.12. Sự phân b c c cl de i A/H5N1 kh c nh theo kh ng gi n (Mà đ : cl de 2.3.4; à x nh l cây: cl de 1; à x nh dương: cl de 7 à
à àng: cl de 2.3.2 (Nguyen, 2010).