Đặc biệt, đã có nhiều người nhiễm và bị tử vong do virus cúm H N1, theo thống kê số người bị nhiễm cúm gia cầm H5N1 của các nước báo cáo với Tổ chức Y tế thế giới (WHO) từ th ng 12 2003 đến 12/4/2012, đã có tới 602 trường hợp mắc cúm H N1, trong số đó 3 trường hợp đã tử vong chiếm tới 58.9%. Indonesia, Việt Nam và Ai Cập là 3 nước có số người tử vong và nhiễm cao nhất do virus cúm A/H5N1trên thế giơi, và đang được Tổ chức Y tế Thế giới-WHO xác định là quốc gia “điểm nóng” có thể xảy ra dịch cúm mới người trong tương lai cần được quan tâm ngăn chặn, do virus cúm A/H5N1có được các điều kiện thuận lợi để tiến hoá thích nghi và lây nhiễm trên người (WHO, 2008).
1.1.2. Bệnh c gi cầ ở Việt N
Bệnh cúm gia cầm xuất hiện lần đầu tiên Việt Nam vào cuối tháng 12/2003 (Bùi Quang Anh, 200 ; BCĐQG, 200 ) do virus cúm gia cầm H5N1 độc lực cao (HPAI) gây ra. Sau đó liên t c tái phát và thường vào lúc chuyển mùa, nhất là v Đông – Xuân. Theo C c Thú y, tính tới năm 2010 thì có 6 đợt dịch (epidemic) cúm gia cầm H5N1 xảy ra:
- Đợt 1: từ tháng 12 2003 đến tháng 3 2004: dịch bệnh đã xảy ra 2.574 xã, phường, 381 huyện, thị thuộc 57 tỉnh, thành phố. Tổng số gia cầm mắc bệnh, chết và tiêu huỷ là 43,9 triệu con (gà: 30,4 triệu; thuỷ cầm: 13,5 triệu). Trong năm 2003 có 3 ca tử vong người do virus cúm H N1.
- Đợt 2: từ tháng 4 đến tháng 11 2004: dịch phát ra rải rác với quy mô nhỏ
các hộ gia đình chăn nuôi gia cầm, bệnh xuất hiện 46 xã, phường tại 32 huyện, quận, thị xã thuộc 17 tỉnh, thành. Thời gian cao điểm nhất là tháng , sau đó giảm dần, đến tháng 11 cả nước chỉ có 1 điểm phát dịch. Tổng số gia cầm mắc bệnh, chết và tiêu huỷ là 84.0 8 con (gà: .999, vịt: 8.132). Trong năm 2004, có 29 ca H5N1 người, có 20 ca tử vong.
- Đợt 3: từ tháng 12 2004 đến tháng 200 : dịch đã xuất hiện 6 0 xã tại 182 huyện thuộc 36 tỉnh, thành phố. Số gia cầm tiêu huỷ là 4 0.49 gà, 82 .689 vịt, ngan. Trong năm 200 , có 61 ca bị nhiễm virus cúm A/H5N1 người, trong đó có 19 ca tử vong.
- Đợt 4: từ tháng 10 200 đến 01/2006: dịch xảy cả 3 miền với 24 tỉnh, thành tái phát. Tổng số gia cầm tiêu huỷ là 3.9 2. 63 con, trong đó, gà: 1.338.523; thuỷ cầm và loài khác: 2.13 .081.
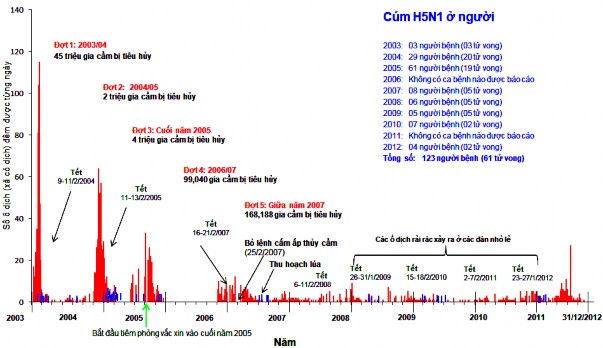
Hình 1.2. Bi đồ bi u diễn dịch c gi cầ do i c A/H5N1 theo thời gi n (Cục Th y, 2012)
- Đợt 5: bắt đầu và kéo dài trong suốt năm 200 . Dịch không tập trung mà rải rác, lẻ tẻ khắp nơi và có thể chia nhiều đợt:
Từ 12 2006 đến 3/2007 dịch xảy ra trên 83 xã, phường của 33 quận, huyện thuộc 11 tỉnh, thành. Tổng số gia cầm mắc bệnh, chết và tiêu huỷ là
103.092 con, trong đó có 13.622 gà ; 89.4 2 ngan, vịt.
Từ 200 đến 8/2007, dịch xảy ra 16 xã, phường của 10 huyện, thị thuộc 23 tỉnh, thành. Tổng số gia cầm mắc bệnh, chết và tiêu huỷ là 294.894 con (21. 2 gà, ; 264. 49 vịt và 8. ngan). Sau khi bị khống chế trong vòng 1 tháng, đến tháng 10 200 , dịch lại tái 1 xã, phường của 9 huyện, quận, thị trấn thuộc 6 tỉnh, thành phố.
Năm 200 có 8 ca bị nhiễm virus cúm A/H5N1 người, trong đó có 5 người chết.
- Đợt 6: từ đầu năm 2008: xảy ra rải rác với 4 đàn gia cầm tại xã, phường của 40 huyện thị thuộc 21 tỉnh phát dịch. Tổng số gia cầm tiêu huỷ là
60.090 con, trong đó có 23.498 gà, 36. 92 thuỷ cầm. Năm 2008 có 6 ca mắc H5N1 người và trong số 6 ca đã tử vong.
Năm 2009, dịch cúm gia cầm đã xảy ra 68 xã, phường, thị trấn của 34 huyện, thị xã thuộc 17 tỉnh, thành với tổng số gia cầm mắc bệnh, chết và tiêu hủy trên 12 .000 con. Năm 2009 có ca mắc H5N1 người và tỷ lệ tử vong là 100% (5/5).
Năm 2010, dịch cúm gia cầm đã xảy ra ít nhất 63 xã, phường của 3 huyện, quận thuộc 24 tỉnh, thành phố, làm hơn 6.000 con gia cầm mắc bệnh, chết và buộc phải tiêu hủy, trong đó chủ yếu là vịt (chiếm hơn 0%). Trong năm 2010, có ca mắc H5N1 người và có 2 ca tử vong (http://www.cucthuy.gov.vn).
1.2. Tình hình nghiên cứ c gi cầ ở Việt N
Bệnh cúm gia cầm xuất hiện Việt nam từ cuối năm 2003 (Trương Văn Dung và Nguyễn Viết Không, 2004). Từ đó đến nay đã có nhiều nghiên cứu về virus và bệnh cúm gia cầm Việt Nam, chủ yếu tập trung nghiên cứu dịch tễ bệnh, khảo sát sự lưu hành của virus, các phương pháp chẩn đoán, nghiên cứu ứng d ng vacxin, xác định hàm lượng kháng thể sau tiêm phòng, điều tra mức độ nhiễm bệnh và giám định phân tử và phân nhóm hệ phả virus gây bệnh, nghiên cứu sản xuất và thử nghiệm vacxin.
Nghiên cứu định type, biến đổi di truyền và gen học tiến hóa của virus cúm A/H5N1 được các cơ quan nghiên cứu của Việt Nam tiến hành ngay từ những tháng đầu tiên xảy ra dịch cúm gia cầm cuối năm 2003. Những chuỗi gen giúp xác định subtype H5, subtype N1 và các gen cấu trúc đã được Viện Công nghệ Sinh học, Viện Pasteur TP Hồ Chí inh, Viện Vệ sinh dịch tễ trung ương, Viện Thú y giải mã và công bố trên Ngân hàng gen (Lê Thanh Hòa, 2006, Dung Nguyen T và cs, 2008). Trên cơ s phân tích trình tự gen kháng nguyên H và N1, các tác giả khẳng định nguồn gốc của virus cúm A gây bệnh trên gia cầm và người tại Việt Nam cùng nhóm với virus A/H5N1 phân lập tại Trung Quốc (Nguyễn Tiến Dũng và cs, 2004; Lê Thanh Hòa, 2006; uramoto và cs, 200 ).
Các biến chủng H N1 của Hong Kong, Trung Quốc phân lập những năm 199 - 2001 và Hàn Quốc, Đài Loan (phân lập năm 2003) đều có nguồn gốc từ chim cút và ngỗng (A Goose Guandong 1 96) vùng Quảng Đông (Trung Quốc), đó là các biến chủng thuộc dòng Quảng Đông (Nguyễn Tiến Dũng và cs, 2004; Lê Thanh Hòa, 2006; Lê Trần Bình và cs, 2006). Như vậy, virus cúm gia cầm gây bệnh gia cầm và người tại Việt Nam là cúm A/H5N1type A thuộc thế hệ mới đã có biến đổi cơ bản về gen H và gen N1, nhưng vẫn có cùng nguồn gốc với H N1 từ vùng địa lý Nam Trung Quốc và Hong Kong (Nguyễn Tiến Dũng và cs, 2004; Lee và cs, 200 ; Simmons và cs, 200 ). Các chủng phân lập những năm 2004- 2006 đã được nghiên cứu khá chi tiết về góc độ gen học và quan hệ phân tử với các chủng trong vùng và thế giới, kết quả khẳng định virus A/H5N1 vùng Nam và Đông Nam Á thuộc nhóm di truyền VT (viết tắt: Vietnam-Thailand-
alaysia), có những đặc tính sinh học nhất định khác với các nhóm vùng Trung Quốc và Hong Kong (Lee và cs, 200 , Chen và cs, 2006). Năm 200 , xuất hiện thêm biến chủng H N1 dưới dòng Phúc Kiến tại Việt Nam, đã và đang làm phức tạp thêm vấn đề dịch tễ học và quan hệ kháng nguyên và miễn dịch, do tỷ lệ tương đồng kháng nguyên HA(H ) và NA(N1) thấp so với các chủng phân dòng Quảng Đông, tuy nhiên vẫn còn có khả năng bảo hộ miễn dịch (Nguyễn ạnh Kiên và cs, 2008, Lê Thanh Hòa và cs, 2008; Nguyễn Tiến Dũng và cs, 2004; Dung Nguyen T và cs, 2008).
Nghiên cứu vấn đề gen học kháng nguyên liên quan đến vacxin và miễn dịch, đã được Viện Công nghệ sinh học, Viện Vệ sinh dịch tễ Trung ương, Viện Pasteur TP Hồ Chí inh, Viện Thú y trung ương tiến hành đó là việc thu thập gen kháng nguyên H và N1 từ các chủng phân lập trên gà, vịt, ngan của Việt Nam các năm 2004 - 2008, và so sánh với trình tự chuỗi gen cúm A/H5N1 của các chủng cường độc đương nhiễm và vacxin của Việt Nam và thế giới (Lê Thanh Hòa, 2004; Nguyễn Tiến Dũng và cs, 2004; Lê Trần Bình và cs, 2006; Lê Thanh Hòa, 2006; Dung Nguyen T và cs, 2008). Năm 200 , xuất hiện thêm chủng H N1 dưới dòng Phúc Kiến (clade 2.3.4) tại Việt Nam, qua phân tích gen giữa các chủng phân lập tại Nghệ An (Việt Nam), A/Dk Vietnam NA114 200 (H N1) và A Dk Vietnam NA 2 200 (H N1), thuộc dòng Phúc Kiến) với các chủng H N1 thuộc phân dòng Quảng Đông, bao
gồm một số chủng làm vacxin đang sử d ng, chỉ đạt 94% (Lê Thanh Hòa và cs, 2008, Nguyễn ạnh Kiên và cs, 2008). Nhận định hỗn hợp virus gây bệnh và phân hóa kháng nguyên của virus cúm A/H5N1 tại Việt Nam cũng đã được xác nhận qua phân tích hàng ch c chủng thu nhận từ nhiều vùng khác nhau trong cả nước (Nguyễn Tiến Dũng và cs, 2004; Dung Nguyên T và cs, 2008). Điều này ảnh hư ng đến dịch tễ, chẩn đoán, phòng trừ và quan hệ lây nhiễm trong tự nhiên (Simmons và cs, 200 ; Alexander, 200 ), cũng như vai trò miễn dịch của các chủng cổ điển đang làm vacxin tại Việt Nam và thế giới (vacxin H N1, chủng gốc: A-Gs-CN-Gd1(96)(H5N1); vacxin H N2, chủng gốc: A-Turkey-ENG- N28(73)(H5N2); vacxin TrovacAIV-H , chủng gốc: A-Tk-IRE- 1378(83)(H5N8)); vacxin H N2, chủng gốc: A-Ck-MEX-Hidalgo- 232(94)(H N2)) và vacxin H N1 thế hệ mới chủng NIB G-14, sử d ng chủng gốc: A Vietnam 1194 2004(H N1) hiện nay đang được Việt Nam nghiên cứu sản xuất (Lê Trần Bình và cs, 2006; Nguyễn Thị Lan Phương và Lê Văn Hiệp, 2006; Lê Trần Bình và cs, 200 ).
Vấn đề chẩn đoán và xây dựng phương pháp phát hiện nhanh và phân biệt cúm A với các tác nhân gây triệu chứng hô hấp khác, cũng như phân biệt các subtype HA và NA đã được các nhà khoa học Việt Nam quan tâm, kết hợp nghiên cứu với các tổ chức thế giới. Phát hiện nhanh H N1 và các subtype khác bao gồm việc sử d ng kháng nguyên hoặc kháng thể, hoặc sinh học phân tử đã được xây dựng thành phương pháp (Chan và cs, 200 ; WHO, 2008). Nghiên cứu vacxin và miễn dịch, các nhà khoa học Việt Nam cũng đã có những đóng góp nhất định về tạo chế phẩm kháng nguyên, tạo vacxin di truyền ngược hoặc vector tái tổ hợp trên nền virus cúm A/H5N1 của Việt Nam (Gao và cs, 2006; Bạch Thị Như Quỳnh, 2006; Lu và cs, 2006; Ge và cs, 200 ).
Về nghiên cứu sản xuất vacxin, Viện Công nghệ sinh học tiến thực hiện đề tài “Nghiên cứu xây dựng qui trình sản xuất vacxin cúm A/H5N1 cho gia cầm” và “Đánh giá chất lượng vacxin phòng bệnh cúm A/H5N1 cho gia cầm tại Việt Nam b ng chủng NIB G-14” trong giai đoạn 2006 - 2008, kết hợp với Viện Thú y, Xí nghiệp thuốc Thú y trung ương (VETVACO), Công ty Thuốc thú y trung ương II (NAVETCO), Trung tâm Kiểm nghiệm thuốc thú y trung ương, thực hiện nghiên cứu có được vacxin sản xuất từ chủng NIB G-14 (Lê Trần Bình,
2007). Đây là chủng vacxin được tạo ra b ng công nghệ di truyền ngược tại Viện Tiêu chuẩn và Kiểm định sinh học Quốc gia (Vương quốc Anh). Kết quả là đã xây dựng được các quy trình sản xuất giống, sản xuất vacxin, kiểm nghiệm và bảo quản vacxin cúm A/H5N1 và kiểm nghiệm miễn dịch đạt chất lượng b ng phương pháp huyết thanh học và thử thách cường độc (Nguyễn Thị Lan Phương và Lê Văn Hiệp, 2006; Lê Trần Bình, 200 ). Hiện nay, vacxin này đã được chuyển giao sản xuất tại NAVETCO với tên thương phẩm là NAVETCO- VIFLUVAC. Song song với những nội dung nghiên cứu về cúm gia cầm gia cầm, các cơ s y tế gồm bệnh viện, viện nghiên cứu (Bệnh viện Nhi trung ương, Viện Vệ sinh dịch tễ Trung ương, Viện Pasteur TP Hồ Chí inh, Viện Vacxin Nha Trang) đều có những triển khai các lĩnh vực nghiên cứu liên quan đến cúm A/H5N1 trên người (Nguyễn Thị Kim Tiến, 200 ; Dinh và cs, 2006; Chan và cs, 200 ; Hatta và cs, 200 ).
1.3. C gi cầ à c c đặc đi m củ i c gi cầm
Cúm gia cầm (Avian Influenza-AI) là một bệnh truyền nhiễm cấp tính của gia cầm, do nhóm virus cúm type A, thuộc họ Orthomyxoviridae gây ra. Đây là nhóm virus có biên độ vật chủ rộng, được phân chia thành nhiều subtype khác nhau dựa trên hai kháng nguyên bề mặt capsid của hạt virus là HA và NA (De Wit, 2008). Nhóm virus cúm A có 16 subtype HA (từ H1 đến H16) và 9 subtype NA (từ N1 đến N9). Sự tổ hợp (reassortment) giữa các subtype HA và NA, về mặt lý thuyết, sẽ tạo ra nhiều subtype khác nhau. ặt khác, virus cúm A có đặc tính quan trọng là dễ dàng đột biến trong gen/hệ gen (đặc biệt gen NA và HA), hoặc trao đổi các gen kháng nguyên với nhau, trong quá trình xâm nhiễm và tồn tại lây truyền giữa các loài vật chủ dẫn đến việc tạo nên nhiều subtype có độc tính và khả năng gây bệnh khác nhau.
Họ Orthomyxoviridae đã được phát hiện bao gồm 4 nhóm virus, đó là: nhóm virus cúm A (Influenza A); nhóm virus cúm B (Influenza B); nhóm virus cúm C (Influenza C); và nhóm Thogotovirus. Các nhóm virus khác nhau b i các kháng nguyên bề mặt capsid, virus cúm A và B là Hemagglutinin (HA), virus
cúm C là Hemagglutinin Esterase Fusion (HEF), và Thogotovirus là Glycoprotein (GP)(Ito và cs, 1998; urphy và Webster, 1996).
1.3.1. Đặc đi inh học phân tử củ i c gi cầ
Hình thái và cấu trúc của virus cúm gia cầm type A đuợc Kawaoka và
urphy (1988) mô tả khá chi tiết. Qua kính hiển vi điện tử, virion có dạng hình khối tròn, hình trứng, hoặc dạng khối dài, đường kính khoảng 80 – 120 nm. Nhiều khi virus có dạng hình sợi dài đến vài µm. Phân tử lượng của hạt virus khoảng 250 triệu Dalton.
B | |
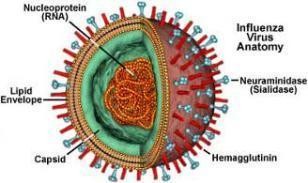
Hình 1.3. Hình ảnh i c A (A)Hình ảnh ph ng; (B Hình th i dưới kính hi n i điện tử (http://micro.magnet.fsu.edu/cells/viruses/influenzavirus.html) |
Có thể bạn quan tâm!
-

 Nghiên cứu một số đặc tính sinh học của vi rút cúm A/H5N1 Clade 7 phân lập ở Việt Nam - 1
Nghiên cứu một số đặc tính sinh học của vi rút cúm A/H5N1 Clade 7 phân lập ở Việt Nam - 1 -
 Nghiên cứu một số đặc tính sinh học của vi rút cúm A/H5N1 Clade 7 phân lập ở Việt Nam - 2
Nghiên cứu một số đặc tính sinh học của vi rút cúm A/H5N1 Clade 7 phân lập ở Việt Nam - 2 -
 M Ph Ng Cấ T C Kh Ng Ng Yên H E L Tinin À Neuraminidase (Www.aht.org.uk)
M Ph Ng Cấ T C Kh Ng Ng Yên H E L Tinin À Neuraminidase (Www.aht.org.uk) -
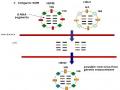 Đặc Đi Tiến Hó À Hình Thành Genotype Củ I C Gi Cầ Gi I Đoạn 1996-2008
Đặc Đi Tiến Hó À Hình Thành Genotype Củ I C Gi Cầ Gi I Đoạn 1996-2008 -
 C C Genotype Củ I C Gi Cầ A/h5N1 Đ C Lực Cao (Li À C , 2007)
C C Genotype Củ I C Gi Cầ A/h5N1 Đ C Lực Cao (Li À C , 2007)
Xem toàn bộ 162 trang tài liệu này.
Vỏ virus có chức năng bao bọc và bảo vệ vật chất di truyền NA của virus, bản chất cấu tạo là màng lipit kép, có nguồn gốc từ màng tế bào nhiễm được đặc hiệu hóa gắn vào các protein màng của virus. Trên bề mặt có khoảng 00 “gai mấu” nhô ra và phân bố dày đặc, mỗi gai mấu dài khoảng 10 - 14 nm có đường kính 4 - 6 nm, đó là những kháng nguyên bề mặt vỏ virus, bản chất cấu tạo là glycoprotein gồm: HA, NA, A (matrix) và các dấu ấn khác của virus (Bender và cs, 1999; Zhao và cs, 2008). Có sự phân bố không đồng đều giữa các phân tử NA và HA (tỉ lệ khoảng 1 NA/4 HA), đây là hai loại protein kháng nguyên có vai trò quan trọng trong quá trình xâm nhiễm của virus tế bào cảm nhiễm (Murphy và Webster, 1996; Uiprasertkul và cs, 200 ).
Hệ gen của virus cúm A là NA sợi đơn âm (viết tắt là (-) ss NA), gồm 8 phân đoạn riêng biệt (HA, NA, , NS, NP, PA, PB1 và PB2) nối với nhau thành một sợi duy nhất bên trong vỏ capsid, mã hóa cho 11 protein tương ứng của virus, trong đó phân đoạn mã hóa cho 2 protein là 1 và 2; phân đoạn NS mã hóa cho 2 protein là NS và NEP, phân đoạn PB1 mã hóa cho 2 protein là PB1 và PB1-F2 (Ito và cs, 1998; Conenello và cs, 200 ) (Hình 1.3.).
- hân đoạn 1 (gen PB2) có kích thước 2431 bp, mã hóa tổng hợp protein enzyme PB2, là tiểu đơn vị thành phần trong phức hợp enzyme polymerase của virus, chịu trách nhiệm kh i đầu phiên mã NA virus. Protein PB2 có khối lượng phân tử theo tính toán khoảng 84.103 Da (trên thực tế là 8 .103 Da) (Murphy và Webster, 1996). Tính thích nghi nhiệt độ cơ thể loài vật chủ được cho là có liên quan đến vị trí amino acid 62 protein PB2 ( virus cúm gia cầm vị trí này là Glu - thích ứng nhiệt độ cơ thể gia cầm khoảng 40oC, còn virus thích nghi trên người là Lys - thích ứng nhiệt độ cơ thể người khoảng 3 oC) (Subbarao và cs, 1998; Wang và cs, 2009).
- hân đoạn 2 (gen PB1) cũng có kích thước 2431 bp, mã hóa tổng hợp enzyme PB1 - tiểu đơn vị xúc tác của phức hợp enzym polymerase trong quá trình tổng hợp NA virus, chịu trách nhiệm gắn mũ NA ( urphy và Webster, 1996). Gần đây, đã có phát hiện thêm một protein (PB1-F2) được mã hóa b i một khung đọc m khác của PB1, có vai trò gây ra hiện tượng apoptosis (hiện tượng tế bào chết theo chương trình) (Tumpey và cs, 2002).
- hân đoạn 3 (gen PA) có kích thước 2233 bp, là phân đoạn gen bảo tồn cao, mã hóa tổng hợp protein enzyme PA có khối lượng phân tử theo tính toán khoảng 83.103 Da (trên thực tế là 96.103 Da). PA là một tiểu đơn vị của polymerase chịu trách nhiệm kéo dài sự phiên mã NA trong quá trình tổng hợp
NA của virus (Luong và Palese, 1992).
- hân đoạn 4 (gen HA) có độ dài thay đổi tuỳ theo từng chủng virus cúm A ( A H1N1 là 1 8 bp; H9N1 là 1 14 bp; H N1 là khoảng 1 04 - 1710 bp). Đây là gen chịu trách nhiệm mã hóa tổng hợp protein HA - kháng nguyên bề mặt virus cúm, gồm hai tiểu phần là HA1 và HA2. Vùng nối giữa HA1 và HA2