Một số virus A/H5N1 thuộc clade được phân lập biên giới phía Bắc và một số chợ gia cầm sống phía Bắc Việt Nam. Sự phân bố của các virus A/H5N1 thuộc các clade khác nhau đã được thu thập và phân tích được trình bày trong bản đồ phân bố (Hình 1.12).
1.3.3.2. Sự hình thành c c genotype
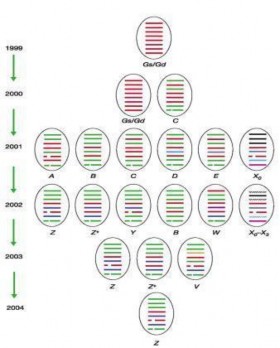
Kể từ khi virus cúm A/H5N1 hiện đại xuất hiện tại Quảng Đông (A Gs CN Gd1 1996(H N1) (Xu và cs, 1999), có tất cả 12 genotype, bao gồm GD, A, B, C, D, E, X(X0-X3), V, Y, W, Z(Z+) và G đã được phát hiện, trong đó các chủng A/H5N1 của Việt Nam là loại mới biến đổi, hầu hết thuộc genotype Z và G Nguyễn Tiến Dũng và cs, 2004; Dung Nguyen T và cs, 2008). Từ năm 2002 tr lại đây, những genotype nguyên thủy của dòng Quảng Đông đã không còn tồn tại (đó là các genotype A, C, D, E), nhưng lại tiến hóa xuất hiện 8 genotype của H N1 (V, W, X1, X2, X3, Y, Z và Z+) (Lee và cs, 200 ; acken và cs, 2006).
Các chủng A H N1 phân lập tại Việt Nam trong những năm 2004 - 2006, thuộc dòng Quảng Đông, tập trung chủ yếu là genotype Z, gần đây xuất hiện thêm genotype G, nhưng được phân biệt làm thành 2 nhóm: nhóm N (North) phổ biến Bắc Việt Nam và nhóm S (South) phân bố miền Nam (Simmons và cs, 200 ; Dung Nguyen T và cs, 2008). Năm 200 , ngoài các chủng và genotype thuộc phân dòng Quảng Đông gây dịch cúm gia cầm, còn có một phân dòng khác đã được phân lập và xác định b ng sinh học phân tử phân tích gen H và N1, đó là dòng Phúc Kiến (Dung Nguyen T và cs, 2008; Lê Thanh Hòa và cs, 2008). Tiến hóa chủng type và dòng tái tổ hợp mới thường định hình và xuất phát từ các tỉnh Nam Trung Quốc (Chen và cs, 2006; Simmons và cs, 200 ; Wang và cs, 2008). Trừ phân dòng Thanh Hải (Qinghai-like sublineage) chưa được phát hiện tại Việt Nam, hai phân dòng Quảng Đông (Guangdong-like sublineage) và Phúc Kiến (Fujian-like sublineage) đã và đang gây dịch cúm gia cầm nước ta, làm cho tình hình dịch tễ và phòng chống có thể tr nên phức tạp trong định hướng đối phó (Dung Nguyen T và cs, 2008).
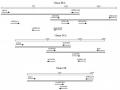
Hình 1.13. C c genotype củ i c gi cầ A/H5N1 đ c lực cao (Li à c , 2007)
Từ năm 2001-200 , đã có 9 genotype (ký hiệu VN) VN1-VN9 xuất hiện hoặc xâm nhập vào Việt Nam (Wan và cs, 2008) được xác định dựa trên phân tích trao đổi chéo các phân đoạn để tái tổ hợp hình ảnh genotype, theo đó dựa trên thành phần H chúng thuộc vào các clade khác nhau.
1.3.4. Tính thích ứng đ ật chủ củ i c A/H5N1
Có thể bạn quan tâm!
-

 Bi Đồ Bi U Diễn Dịch C Gi Cầ Do I C A/h5N1 Theo Thời Gi N (Cục Th Y, 2012)
Bi Đồ Bi U Diễn Dịch C Gi Cầ Do I C A/h5N1 Theo Thời Gi N (Cục Th Y, 2012) -
 M Ph Ng Cấ T C Kh Ng Ng Yên H E L Tinin À Neuraminidase (Www.aht.org.uk)
M Ph Ng Cấ T C Kh Ng Ng Yên H E L Tinin À Neuraminidase (Www.aht.org.uk) -
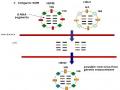 Đặc Đi Tiến Hó À Hình Thành Genotype Củ I C Gi Cầ Gi I Đoạn 1996-2008
Đặc Đi Tiến Hó À Hình Thành Genotype Củ I C Gi Cầ Gi I Đoạn 1996-2008 -
 Kết Quả Tiê Phòng Vacxin C Gi Cầ Chương T Ình Q C Gia
Kết Quả Tiê Phòng Vacxin C Gi Cầ Chương T Ình Q C Gia -
 Bảng Tổng Hợp Liệ Tính To N Theo Phương Ph P Reed-Muench
Bảng Tổng Hợp Liệ Tính To N Theo Phương Ph P Reed-Muench -
 Ch Ẩn Bị Ix (Hỗn Hợp Phản Ứng Rt- Cr Giải T Ình Tự Gen
Ch Ẩn Bị Ix (Hỗn Hợp Phản Ứng Rt- Cr Giải T Ình Tự Gen
Xem toàn bộ 162 trang tài liệu này.
Vật chủ tự nhiên của tất cả các chủng virus cúm A/H5N1 là chim hoang dã (chủ yếu là vịt trời), đây là nguyên nhân lan truyền virus trong tự nhiên rất khó kiểm soát. Virus cúm A có khả năng gia tăng biên độ vật chủ của chúng trong quá trình lây truyền tự nhiên (Kash và cs, 2006; De Wit, 2008). Nhờ đặc tính luôn thay đổi kháng nguyên trong tự nhiên, virus cúm A có khả năng xâm nhiễm nhiều loài vật chủ trung gian khác nhau như gia cầm, một số loài động vật có vú (hải cẩu, cá voi, ngựa, lợn) và cả người, tạo nên tính thích ứng lan truyền “nội loài” như gà - gà, hay “ngoại loài” như gà - lợn; gà - lợn - người (Hình 1.1 ). Vịt (vịt trời) và một số loài thuỷ cầm khác (ngỗng) luôn luôn là vật chủ tàng trữ nguồn virus gây nhiễm (Webster, 1998; Webster và cs, 2002; Chen và cs, 2004; Hulse-Post và cs, 200 ). Đặc điểm thích ứng vật chủ này là điều
kiện thuận lợi cho virus cúm A trao đổi, tái tổ hợp các phân đoạn gen, đặc biệt là các phân đoạn gen kháng nguyên (gen “độc” HA và NA) giữa các chủng, tạo ra chủng virus cúm mới có khả năng thích ứng xâm nhiễm loài vật chủ mới của chúng đặc biệt khi chúng vượt qua được “rào cản loài” dễ dàng thích ứng lây nhiễm gây bệnh từ gia cầm sang người và giữa người với người (Horimoto và Kawaoka, 1994; Hilleman, 2002; Chen và cs, 2004). Trong lịch sử các đại dịch cúm người, lợn thường là vật chủ trung gian chuyển tiếp giúp cho virus cúm A biến đổi để dễ dàng lây nhiễm sang người gây nên bệnh dịch (Ito và cs, 1998; Zhu và cs, 2008). Ví d , cúm A H3N2 là kết quả tái tổ hợp tự nhiên của virus cúm A H2N2 của người và virus chứa gen H3 trong tự nhiên thông qua đồng nhiễm trên lợn, gây nên đại dịch cúm châu Á năm 1968 (Nicholson và cs, 2003; Wanasawaeng và cs, 2009; Horimoto và Kawaoka, 1994).
Hình 1.14. M i q n hệ lây nhiễ à thích ứng c c loài ật chủ củ i c A (Wahlgren, 2011)
1.3.5. Cơ chế xâ nhiễ gây bệnh củ i c A t ong tế bào ật chủ
Virus cúm A/H5N1 kí sinh nội bào bắt buộc, quá trình xâm nhiễm và nhân lên của virus xảy ra chủ yếu các tế bào biểu mô đường hô hấp, đường tiêu hóa
của cơ thể nhiễm ( urphy và Webster, 1996; Nicholson và cs, 2003), có những nét đặc trưng như sau:
- Quá trình xâm nhiễm của virus cúm A được m đầu b ng sự kết hợp của HA và th thể thích ứng của nó trên bề mặt các tế bào này, và cuối cùng là giải phóng hệ gen của virus vào trong bào tương của tế bào nhiễm (Hình 1.15).
- Quá trình nhân lên của NA virus cúm A chỉ xảy ra trong nhân của tế bào, đây là đặc điểm khác biệt so với các virus khác (quá trình này xảy ra trong nguyên sinh chất), và cuối cùng là giải phóng các hạt virus ra khỏi tế bào nhiễm nhờ vai trò của enzyme neuraminidase. Thời gian một chu trình xâm nhiễm và giải phóng các hạt virus mới của virus cúm chỉ khoảng vài giờ (trung bình 6 h). Sự tạo thành các hạt virus mới không phá tan tế bào nhiễm, nhưng các tế bào này bị rối loạn hệ thống tổng hợp các đại phân tử, và rơi vào quá trình chết theo chương trình (apoptosis) làm tổn thương mô của cơ thể vật chủ (Wanasawaeng và cs, 2009; Tumpey và cs, 2002).
- Sau khi được giải phóng vào trong bào tương tế bào nhiễm, hệ gen của virus sử d ng bộ máy sinh học của tế bào tổng hợp các protein của virus và các
NA vận chuyển ph thuộc NA ( NA-dependent NA transcription). Phức hợp protein – NA của virus được vận chuyển vào trong nhân tế bào (Beard, 1998).
- Trong nhân tế bào các NA hệ gen của virus tổng hợp nên các sợi dương từ khuôn là sợi âm của hệ gen virus, từ các sợi dương này chúng tổng hợp nên
NA hệ gen của virus mới nhờ NA-polymerase. Các sợi này không được Adenine hóa (gắn thêm các Adenine - gắn mũ) đầu ’- và 3’-, chúng kết hợp với nucleoprotein (NP) tạo thành phức hợp ribonucleoprotein ( NP) hoàn chỉnh và được vận chuyển ra bào tương tế bào. Đồng thời, các NA thông tin của virus cũng sao chép nhờ hệ thống enzyme từng phân đoạn gen của virus, và được enzyme PB2 gắn thêm 10 - 12 nucleotide Adenin đầu ’-, sau đó được vận
chuyển ra bào tương và dịch mã tại lưới nội bào có hạt để tổng hợp nên các protein của virus (Hình 1.1 ).
Hình 1.15. M hình cơ chế xâ nhiễ à nhân lên củ i c A ở tế bào chủ (Beard, 1998).
- Các phân tử NA và HA của virus sau khi tổng hợp được vận chuyển gắn lên mặt ngoài của màng tế bào nhiễm nhờ bộ máy Golgi, gọi là hiện tượng “nảy chồi” của virus. NP sau khi tổng hợp được vận chuyển tr lại nhân tế bào để kết hợp với NA thành NP của virus. Sau cùng các NP của virus được hợp nhất với vùng “nảy chồi”, tạo thành các “chồi” virus gắn chặt vào màng tế bào chủ b i liên kết giữa HA với th thể chứa sialic acid. Các NA phân cắt các liên kết này và giải phóng các hạt virus trư ng thành tiếp t c xâm nhiễm các tế bào khác ( urphy và Webster, 1996; Nayak và cs, 2004).
1.3.6. Đ c lực à khả năng gây bệnh củ i c gi cầm
Nghiên cứu mức độ phân tử cho thấy, khả năng lây nhiễm virus ph thuộc vào tác động của men protease của vật chủ đến sự phá vỡ các liên kết hoá học sau khi dịch mã của phân tử liên kết. Tính th cảm của ngưng kết tố và sự phá vỡ liên kết của enzyme protease lại ph thuộc vào số lượng các axit amin polybasic tại điểm bắt đầu phá vỡ liên kết. Các enzym giống như trypsin có khả
năng phá vỡ liên kết khi chỉ có một phân tử arginin, trong khi đó các men protease khác lại cần nhiều axit amin polybasic, vì thế một số nước đánh giá độc lực của virus trên cơ s gây nhiễm cho gia cầm và sau đó phân tích sự sắp xếp các axit amin của virus. C thể người ta quan tâm đến việc giải trình tự vùng “cleavage site” (chuỗi nối) của gen HA mà sự có mặt của các aminoaxit như arginine (R) hay lysine (K) sẽ cho phép dự đoán r ng virus đó có độc lực cao hay không (Hình 1.16). Tuy nhiên người ta thường chỉ thực hiện điều này đối với các virus thuộc subtype H và H .
Trong thực tế người ta chia virus cúm ra làm 2 loại: Loại virus có độc lực thấp – LPAI và loại virus có độc lực cao HPAI.
- LPAI là loại virus khi phát triển trong cơ thể nhiễm, có thể gây bệnh cúm nhẹ không có triệu chứng lâm sàng điển hình và không làm chết vật chủ. Đây là loại virus lây truyền rộng rãi và tạo nên các ổ bệnh trong tự nhiên của virus cúm
A. Loại này có thể trao đổi gen với các chủng virus có độc lực cao đồng nhiễm trên cùng một tế bào, và tr thành một loại virus HPAI nguy hiểm.
- HPAI là loại virus cúm A có khả năng gây tổn thương nhiều cơ quan nội tạng trong cơ thể nhiễm, trên gia cầm chúng thường gây chết 100% số gia cầm bị nhiễm trong vòng 48-72 giờ sau nhiễm. Virus loại HPAI phát triển tốt trên tế bào xơ phôi gà, tế bào thận chó ( DCK) không có trypsin. Các v dịch lớn đều do virus HPAI gây ra, thường là virus có kháng nguyên H và H .
Hình 1.16. Minh hoạ ùng “cle ge ite” củ i c H5N1 đ c lực thấp (L AI à đ c lực cao (H AI (W gne à c , 2002
Virus cúm A có tính thích ứng lây nhiễm cao với biểu mô đường hô hấp, gây bệnh chủ yếu đường hô hấp, và cũng có thể tác động gây tổn thương nhiều cơ quan khác trong cơ thể của các động vật cảm nhiễm, do đó còn được gọi là virus hướng đa phủ tạng (Wanasawaeng và cs, 2009; Nicholson và cs, 2003). Khả năng gây bệnh của virus cúm A ph thuộc vào độc lực và tính thích nghi vật chủ của từng chủng virus. Thông thường chúng không gây bệnh hoặc chỉ gây bệnh nhẹ giới hạn đường hô hấp của chim hoang dã và gia cầm nhiễm, nhưng một số chủng cường độc (H , H và H1, H2 và H3) có thể gây nên bệnh nặng hầu hết các cơ quan trong cơ thể, gây nên dịch cúm gia cầm và người có lẽ do tính thích ứng th thể sialic của chúng (Subbarao và Joseph, 200 ; Xu và cs, 1999). Hầu hết các chủng virus cúm A nhân lên rất tốt trong phôi gà sau lần cấy truyền thứ nhất, tuy nhiên các chủng cường độc subtype H và H gây chết phôi gà ngay sau vài giờ, cả khi hàm lượng virus rất thấp chưa được nhân lên nhiều, và có thể gây bệnh cúm thực nghiệm trên chuột lang, chuột hamster, chồn đất (Horimoto và Kawaoka, 1994, De Wit, 2008).
Sau bị khi nhiễm virus cúm A, cơ thể vật chủ sinh ra đáp ứng miễn dịch chống lại virus bảo vệ cơ thể, nhưng đáp ứng miễn dịch này có thể không có tác d ng bảo vệ hoàn toàn cho những lần nhiễm sau, do virus cúm A luôn có sự biến đổi kháng nguyên của nó trong quá trình lưu hành tự nhiên, và không có đáp ứng miễn dịch chéo giữa các chủng virus cúm A (Wanasawaeng và cs, 2009). Do đó, khi xuất hiện những biến chủng virus cúm A có đặc tính kháng nguyên khác với các chủng virus trước đó, cơ thể nhiễm sẽ không hoặc ít có đáp ứng miễn dịch bảo hộ thích ứng với chủng virus cúm mới. Đây là nguyên nhân làm cho gia cầm và con người thường bị mắc bệnh cúm nhiều lần trong năm, và các đợt dịch cúm xảy ra về sau thường nặng nề hơn và có thể gây nên đại dịch cúm mới (Doherty và cs, 2006).
1.3.7. Triệu chứng lâ àng của gia cầm mắc bệnh c
Virus cúm gia cầm độc lực cao thường gây bệnh rất trầm trọng cho gia cầm và thông thường tỷ lệ tử vong cao đặc biệt là với gia cầm cạn. Các loài thủy cẩm, dã cầm và một số loài lông vũ khác có thể có độ mẫn cảm thấp hơn, nhưng có thể tr thành động vật mang trùng
1.2.7.1. Đặc đi t iệ chứng t ên gà
- Chết đột ngột
- Ủ rũ, lông xơ xác, bỏ ăn
- Giảm năng suất đẻ trứng
- Phù thũng đầu và cổ
- ào và tích sưng to hoặc có màu tím tái
- Xuất huyết lấm chấm trên mặt các màng tương
- Khát nước nghiêm trọng
- Tiêu chảy ra nước loãng màu xanh ban đầu, sau đó chuyển sang trong hoặc màu trắng
- àng kết mạc sưng hoặc t máu đôi khi có xuất huyết
- Xuất huyết da chân
- Triệu chứng hô hấp tùy thuộc và mức độ thương tổn trong khí quản
- Chảy nước mũi, miệng
- Con vật yếu sức,
- Ho, hắt hơi
- Triệu chứng thần kinh, loạng choạng
- Ngừng đẻ, trứng dị hình trước khi ngừng đẻ
- Tỷ lệ chết có thể tới 100%, có thể chết trước khi có triệu chứng
1.2.7.2. Đặc đi t iệ chứng t ên thủy cầ
- Ủ rũ
- Bỏ ăn, tiêu chảy
- Triệu chứng thần kinh, quay cuồng, co giật
- ắt màu khói, đ c
- Năng suất trứng giảm
- Có thể chết đột ngột
- Những con khỏi bệnh yếu ớt, có thể đẻ trứng tr lại vài tuần sau khi khỏi
bệnh
- Tỷ lệ tử vong có thể tới 100%