và đang được sử dụng làm nhân tố đấu tranh sinh học hiệu quả chống lại các loài côn trùng gây hại mùa màng, cây rừng. Một số khác được sử dụng trong các quy trình công nghệ sinh học hoặc trong y học [102].
Theo Evans và cộng sự, nấm ký sinh côn trùng được chia thành 4 nhóm: Nhóm ký sinh trong các nội quan, khoang cơ thể ký chủ; nhóm phát triển trên lớp cuticun vỏ cơ thể của ký chủ; nhóm mọc trên ký chủ và nhóm cộng sinh trên ký chủ [52]. Bào tử nấm tiếp xúc với bề mặt ký chủ, khi gặp điều kiện thuận lợi sẽ mọc mầm, xâm nhiễm vào bên trong cơ thể ký chủ qua lớp vỏ cuticun. Ngoài ta, nấm ký sinh côn trùng có thể xâm nhiễm vào cơ thể côn trùng qua con đường hô hấp, tiêu hóa. Nấm mọc khắp bên trong cơ thể côn trùng và sản xuất các độc tố nhằm gây chết côn trùng và ngăn chặn sự cạnh tranh của vi sinh vật khác. Chu trình xâm nhiễm của nấm ký sinh côn trùng bao gồm ba giai đoạn chính: Giai đoạn xâm nhập (bào tử nấm mọc mầm, giải phóng các enzyme ngoại bào phân hủy lớp vỏ cuticun côn trùng xâm nhập vào trong xoang cơ thể côn trùng); giai đoạn phát triển trong cơ thể côn trùng (nấm tiếp tục phát triển hình thành các sợi nấm phát tán khắp bên trong cơ thể côn trùng cho đến khi côn trùng chết); giai đoạn phát triển sau khi ký chủ chết (nấm sử dụng cạn kiệt nguồn dinh dưỡng bên trong cơ thể côn trùng chuyển sang giai đoạn hình thành bào tử) [174].
Cho đến nay, 65 loài Chytridiomycota, 474 loài Zygomycota, 238 loài Basidiomycota và gần 1000 loài Ascomycota đã được ghi nhận [19, 222]. Trong các nghiên cứu đã công bố, những loài nấm túi thuộc chi Metarhizium(M. anisopliae, M. robertsii, M. brunneum, M. lepidiotae, M. globosum, M.acridum, M. majus, M. flavoviride, M. rileyi, M. pingshaense, M. lepidiotaevà M. guizhouense), chi Beauveria (B. bassiana và B. brongniartii), chi Isaria(I. fumosorosea, I. farinosa và I. tennuipes), chi Ophiocordyceps (O. sinensis,O. unilateralis), chi Cordyceps (C. militaris), chi Torubiella (T. ratticaudata),
chi Pochonia (P. chlamydosporia), chi Lecanicillium (L. lecani, L.longisporum), chi Hirsutella (H. thompsonii, H. nodulosa, H. aphidis), và cácloài Paecilomyces variotii, Purpureocillium lilacinum được nghiên cứu vàứng dụng phổ biến nhất [19, 86, 127]. Mặc dù các loài thuộc Entomophthorales (ví dụ như Furia, Conidiobolus, Entomophaga, hoặc Erynia) có khả năng diệt côn trùng cao nhất nhưng do đặc điểm sinh học của chúng (khó khăn trong nuôi cấy và phát triển sản phẩm) nên chúng không được sử dụng làm thành phần của chế phẩm sinh học [109].
Nấm ký sinh côn trùng có vai trò quan trọng đối với hệ sinh thái. Ví dụ, các loài thuộc chi Metarhizium và Beauveria, thường được tìm thấy trong đất, không chỉ kiểm soát các quần thể côn trùng tự nhiên mà còn sống nội cộng sinh với thực vật. Một số loài nấm ký sinh côn trùng sống nội cộng sinh ở rễ, thân và lá thực vật [77]. Ví dụ như loài M. robertsii và B. bassiana cung cấp nitơ được đồng hóa cho cây trồng trong quá trình ký sinh của côn trùng, do đó hỗ trợ sự phát triển của cây trồng [143]. Beauveria bassiana được ghi nhận sống nội cộng sinh trong khoảng 25 loài thực vật, góp phần kiểm soát sâu bệnh và nấm bệnh thực vật [186]. Beauveria bassiana còn sống trên lá và chồi bên cạnh rễ cây làm cho cây có khả năng chống lại côn trùng tốt hơn [140], đồng thời cũng bảo vệ thành công thực vật khỏi mầm bệnh vi sinh vật bằng cách ngăn chặn các tác nhân gây bệnh hoặc tăng phản ứng phòng vệ của thực vật [119]. Tương tự, Lecanicillium cũng ngăn ngừa bệnh nấm bằng cách phát triển trên bề mặt của lá, hạn chế các chất dinh dưỡng có sẵn và tạo ra các hợp chất kháng khuẩn ngoài việc gây ra các phản ứng của cây trong khi xâm nhập vào rễ cây [119]. Trong số nấm ký sinh côn trùng, I. fumosorosea dường như tương tác mạnh với thực vật. Điều này có lẽ là do Isaria spp. nhạy cảm với các chất hóa học do thực vật tiết ra và thuộc hệ thống phòng thủ chống lại các mầm bệnh thực vật [220]. Hiện nay nấm ký sinh côn trùng được sử dụng
phổ biến nhất để kiểm soát sinh học đối với côn trùng gây hại trong phòng thí nghiệm, nhà kính hoặc trên đồng ruộng.
1.1.2.2. Các hợp chất trao đổi thứ cấp từ nấm ký sinh côn trùng
Nấm ký sinh côn trùng có khả năng sinh tổng hợp các hợp chất hữu cơ có trọng lượng phân tử thấp được gọi là chất chuyển hóa thứ cấp. Các hợp chất này cần thiết cho lây nhiễm mầm bệnh một cách hiệu quả bằng cách làm tổn thương hệ thần kinh hoặc giảm sức đề kháng của côn trùng [48].
Các chất chuyển hóa thứ cấp được xác định bằng phương pháp sắc ký kết hợp với kỹ thuật khối phổ và sinh học phân tử. Dựa trên cấu trúc hóa học, chúng có thể được chia thành các nhóm sau: Cyclic depsipeptide, peptide, dẫn xuất amino acid, polyketide, peptide hybrid, và terpenoid [48], [196]. Các chất chuyển hóa từ nấm ký sinh côn trùng rất đa dạng và có phổ hoạt tính mạnh đã và đang thu hút được sự quan tâm của các nhà nghiên cứu.
Đơn cử như cordycepin, thuộc nhóm octadepsipeptide, là sản phẩm của quá trình chuyển hóa thứ cấp của nấm ký sinh côn trùng Cordyceps militaris và Isaria cicadae. Cordycepin đã được sử dụng trong y học Trung Quốc trong nhiều năm vì đặc tính chống oxy hóa, chống viêm, kháng u và bảo vệ thần kinh [196]. Ngoài ra cordycepin được chứng minh làm giảm các triệu chứng viêm xương khớp, tức là viêm bao hoạt dịch, bệnh lý khớp và giảm đau [20]. Cordycepin còn được chứng minh khả năng ức chế sự già đi của tế bào ở loài gặm nhấm thông qua con đường NRF2 (yếu tố nhân erythroid 2 liên quan đến yếu tố 2) và AMPK (AMP - hoạt hóa protein kinase) trong quá trình viêm. Ứng dụng này mang lại hy vọng cho tương lai để ngăn chặn quá trình lão hóa tế bào và loét bức xạ [196].
Các chất chuyển hóa thứ cấp của nấm ký sinh côn trùng là một tập hợp các hợp chất có hoạt tính đa dạng. Vì vậy, các nhà khoa học hiện nay đang
nghiên cứu sâu hơn về cấu trúc, đặc tính của các hợp chất này; tìm kiếm các hợp chất mới và tìm hiểu con đường sinh tổng hợp.
1.2. Nấm ký sinh côn trùng sinh tổng hợp cyclooligomer depsipeptide
1.2.1. Cyclooligomer depsipeptide
1.2.1.1. Cấu trúc hóa học
Cyclooligomer depsipeptide (COD) là các nonribosomal peptide đặc biệt thuộc các nhóm hợp chất peptolide mạch vòng. Đây là những peptide có phổ hoạt tính sinh học rộng được sản sinh từ các vi sinh vật thông qua con đường sinh tổng hợp peptide ngoài ribosome bởi các enzyme phức hợp chứa một hay nhiều protein lớn (NRPSs) [47, 158].
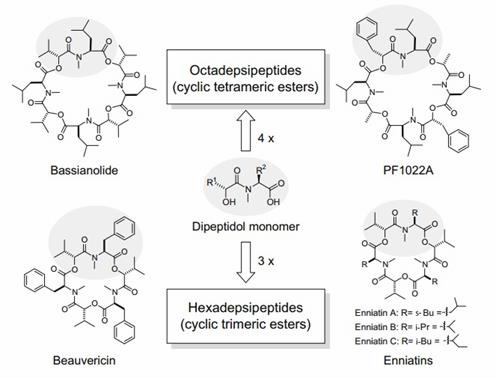
Hình 1.1. Cấu trúc một số loại cyclooligomer depsipeptide [204]
Những nghiên cứu về các COD này bắt đầu sớm nhất là vào những năm 1940, tiêu biểu như tách chiết enniatin A từ nấm Fusarium orthocera var. enniatinum [57]. Tuy nhiên, thuật ngữ “cyclooligomer depsipeptide” còn được gọi là “cyclooligomeric depsipeptide” lần đầu tiên được giới thiệu trong các tài liệu khoa học vào năm 2008 [205].
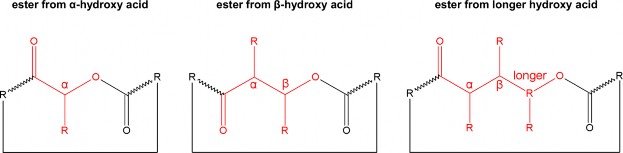
Hình 1.2. Cấu trúc chung của các liên kết este giữa nhóm carboxyl C- terminus α-hydroxy acid, ß-hydroxy acid và hydroxy- acid chuỗi dài [173]
COD dùng để mô tả các peptide vòng, trong đó mỗi vòng là các amino acid và các hydroxy-acid liên kết với nhau bằng các liên kết amide và liên kết este [17, 101, 121]. Thông thường cyclooligomer depsipeptide gồm 2, 3 hoặc
4 đơn phân. Trong đó mỗi đơn phân được cấu tạo bởi ít nhất một 2- hydroxycarboxylic acid và một 2-amino acid [169] (Hình 1.1).
Theo khóa phân loại của Taevernier và cộng sự, các peptolide tuần hoàn khác nhau bởi loại este hay sự hình thành liên kết este giữa nhóm carboxyl C-terminus và nhóm hydroxy-acid cụ thể (α-hydroxy acid, ß- hydroxy acid hoặc hydroxy-acid chuỗi dài) và số lượng liên kết este trong cấu trúc vòng. Các COD như bassianolide, beauvericin, enniatin và PF1022A được nhóm lại vào cùng một lớp (mã lớp 121) sau 3 lần phân tách [173] (Hình 1.2). Theo đó cyclooligomer depsipeptide bao gồm các peptolides mà trong cấu trúc mạch vòng của chúng có 2 hay nhiều liên kết este được hình thành giữa nhóm carboxyl với nhóm α-hydroxy acid, các liên kết este trong vòng được sắp xếp một cách xen kẽ, đều đặn tạo nên cấu trúc đối xứng [169].
1.2.1.2. Các dạng cyclooligomer depsipeptide từ nấm
Hiện nay các depsipeptide từ nấm được tìm thấy rất đa dạng và phong phú. Tuy nhiên trên thực tế chỉ có 1348 depsipeptide được mô tả cấu trúc rõ ràng, đã và đang được các nhà khoa học nghiên cứu và sử dụng. Trong số đó có khoảng hơn 80 COD (Bảng 1.1) [173].
Bảng 1.1. Một số cyclooligomer depsipeptide (COD) phổ biến ở nấm
COD | Nấm | Tham khảo | |
1 | Allobeauvericina-n | Paecilomyces tenuipes BCC1614 | [126] |
2 | Bassianolide | Beauveria bassiana, Verticillium lecanii | [84], [122] |
3 | Beauvericina-n | Aspergillus terreus GX7-3B, Beauveria bassiana ATCC7159, Cordyceps cicadae, Fusarium redolens… | [44], [206], [193], [202] |
4 | Cardinalisamidea-n | Cordyceps cardinalis | [180] |
5 | Cordycecin A | Cordyceps cicadae | [193] |
6 | Enniatinsa-n | Fusarium tricinctum, Acremonium sp. BCC2629, Verticillium hemipterigenum BCC1449, Halosarpheia sp. 732… | [211], [191], [167], [96] |
7 | Hirsutellide A | Hirsutella kobayasii | [190] |
8 | PF1022a-n | Rosellina sp. PF1022, Mycelia sterilia PF1022, Trichoderma asperellum | [150], [129], [45] |
9 | Pseudoxylallemycina- n | Pseudoxylaria sp. X802 | [61] |
10 | Verticilidea-n | Verticillium sp. FKI2679 | [128] |
Có thể bạn quan tâm!
-

 Nghiên cứu đa dạng và sinh tổng hợp Cyclooligomer depsipeptide của nấm ký sinh côn trùng tại Khu Bảo tồn thiên nhiên Copia và Vườn quốc gia Xuân Sơn - 1
Nghiên cứu đa dạng và sinh tổng hợp Cyclooligomer depsipeptide của nấm ký sinh côn trùng tại Khu Bảo tồn thiên nhiên Copia và Vườn quốc gia Xuân Sơn - 1 -
 Nghiên cứu đa dạng và sinh tổng hợp Cyclooligomer depsipeptide của nấm ký sinh côn trùng tại Khu Bảo tồn thiên nhiên Copia và Vườn quốc gia Xuân Sơn - 2
Nghiên cứu đa dạng và sinh tổng hợp Cyclooligomer depsipeptide của nấm ký sinh côn trùng tại Khu Bảo tồn thiên nhiên Copia và Vườn quốc gia Xuân Sơn - 2 -
 Khả Năng Ức Chế Sự Sinh Trưởng Của Tế Bào Ung Thư Của Một Số Cod
Khả Năng Ức Chế Sự Sinh Trưởng Của Tế Bào Ung Thư Của Một Số Cod -
 Nấm Cordyceps Spp. Sinh Tổng Hợp Cyclooligomer Depsipeptide Cordyceps Là Một Chi Nấm Thuộc Họ Cordycipitaceae, Bộ Hypocreales,
Nấm Cordyceps Spp. Sinh Tổng Hợp Cyclooligomer Depsipeptide Cordyceps Là Một Chi Nấm Thuộc Họ Cordycipitaceae, Bộ Hypocreales, -
 Tách Chiết, Tinh Sạch Và Nghiên Cứu Cấu Trúc Cyclooligomer Depsipeptide Từ Nấm Ký Sinh Côn Trùng
Tách Chiết, Tinh Sạch Và Nghiên Cứu Cấu Trúc Cyclooligomer Depsipeptide Từ Nấm Ký Sinh Côn Trùng
Xem toàn bộ 228 trang tài liệu này.
Dựa vào số lượng các amino acid và các hydroxy-acid trong hợp chất có thể chia COD từ nấm thành 4 loại chính bao gồm cyclotetrapeptide, cyclohexadepsipeptide, cyclooctadepsipeptide và diketomorpholine [169, 196]: Cyclotetrapeptide bao gồm 6 hợp chất là pseudoxylallemycin (A-F) được phân lập từ chi nấm Pseudoxylaria [216]. Pseudoxylallemycin (A-F) là các peptide mạch vòng có kích thước trung bình chứa các amino acids thơm, kỵ nước được xếp xen kẽ với các nhóm N-methyl. Ví dụ như Pseudoxylallemycin A là một dimer mạch vòng của monomer N-methyl-L- leucine-L-phenylalanine. Pseudoxylallemycin B là dimer mạch vòng của monomer N-methyl-Lleucine-4-(buta-2,3-dienyloxy)-L-phenylalanine. Riêng
Pseudoxylallemycin B, C, D còn sở hữu một gốc allenyl hiếm gặp… [61].
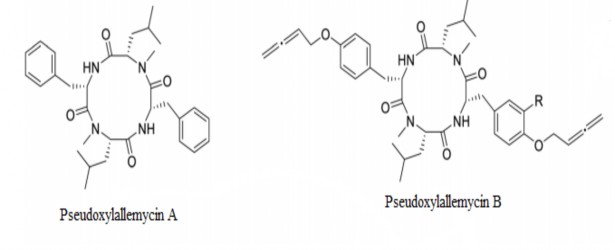
Hình 1.3. Cấu trúc Pseudoxylallemycin [216]
Cyclohexadepsipeptide là nhóm COD lớn nhất, được tìm thấy phổ biến ở các chi nấm Acremonium, Aspergillus, Beauveria, Cordyceps, Fusarium, Isaria, Nigrospora, Peacilomyces, và Verticillium [196]. Các COD chính thuộc nhóm cyclohexadepsipeptide bao gồm beauvericins, beauvericin (A, B, C, D, F, J, G1, G2, G3, H1, H2, H3), allobeauvericin (A, B, C), enniatin (A, A1, A2, B1, B2, B3, B4, C, E1, E2, F, G, H, I, J1, J2, J3, K1, L, M1, M2, MK1688, N, O1, O2, O3, P1, P2, Q, R, S, T, U, V), hirsutellide A,
cardinalisamide (A, B, C), cordycecin A, verticilide B1…[79, 98, 134, 171]. Beauvericin là một trimer mạch vòng của dipeptidol. Trong đó mỗi dipeptidol được hình thành từ (2R) -2-hydroxy-3-methylbutanoic acid (D- hydroxyisovaleric acid, D-Hiv) và N-methyl-L-phenylalanine (N-Me-Phe) [199]. Enniatin có cấu trúc trimeric tương tự như beauvericin. Tuy nhiên, các gốc N-Me-Phe thơm đặc trưng trong các đơn phân beauvericin dipeptidol được thay thế bằng aliphatic NMe-Ile (enniatin A), N-Me-Val (enniatin B) hoặc N-Me-Leu (enniatin C) [39, 104] (Hình 1.1). Khác với beauvericin và enniatin, hirsutellide A là một dimer tuần hoàn của tripeptidolmonomer D-2- hydroxy-3-phenylpropanoic acid-L-allo-isoleucine–N-methylglycine [190]…
Cyclooctadepsipeptide bao gồm các COD mà bên trong cấu trúc có chứa 8 amino acid và hydroxy-acid. Tiêu biểu như các bassianolide được phân lập từ nấm Beauveria bassiana, Verticillium lecanii hay một số loài thuộc chi Xylaria. Vòng macrolactone trong cấu trúc của bassianolide được hình thành dưới dạng tetramer của dipeptidol monomer D-Hiv-N-Me-Leu. Các monomer của bassianolide tương tự với monomer của cyclohexadepsipeptide enniatin C [84, 204]. Một nhóm các cyclooctadepsipeptide phổ biến khác là PF1022 (A, B, C, D, E, F, G, H). Cấu trúc của PF1022 được cấu tạo từ hydroxycarboxylic acid D-Lac hoặc D- PheLac và amino acid N-Me-Leu liên kết với nhau hình thành lên các dipeptidol. Trong quá trình sinh tổng hợp PF1022, từ các dipeptidol trải qua quá trình cyclotetramer hóa hình thành nên các tetradepsipeptide. Mỗi PF1022 bao gồm 2 tetradepsipeptide sắp xếp đối xứng nhau [198] (Hình 1.1). Trong tự nhiên PF1022 được ghi nhận tìm thấy ở chi nấm Rosellinia [115]. Ngoài ra, các cyclooctadepsipeptide điển hình khác như verticilide (A1, A2, A3) [117], glomosporin [151],…
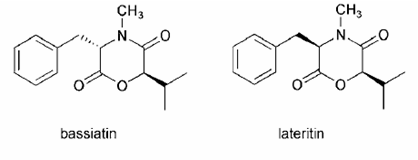
Hình 1.4. Cấu trúc một số diketomorpholine [169]
Diketomorpholine là các COD mà trong cấu trúc chỉ gồm một monomer duy nhất. Điển hình như bassiatin và lateritin - diketomorpholine được tìm thấy ở một số chi nấm Beauveria, Cordyceps, Fusarium, Gibberella hay Isaria [22, 68, 83, 137]. Bassiatin và lateritin là hai cấu trạng diketomorpholine với nhau, được cấu tạo từ (2R) -2-hydroxy-3-