men của chủng Verticillium sp. FKI-2679 cũng được phân lập ghi nhận có thể tạo ra 1 loạt các COD có khả năng ức chế acyl-CoA: cholesterol acyltransferase (ACAT1 và ACAT2) là verticilide A1, A2, A3 [128].
Khả năng tạo ra COD và các hoạt chất sinh học khác của các loài thuộc chi Paecilomyces đã được báo cáo trong nhiều nghiên cứu. Điển hình như khi nghiên cứu chủng nấm gây bệnh côn trùngPaecilomyces tenuipes BCC 1614, Nilanonta đã phân lập được và xác định được hai cyclohexadepsipeptide có khả năng kháng khuẩn là beauvericin và beauvericin A [125]. Cùng tác giả này với nghiên cứu tiếp theo vào năm 2002, đã phát hiện thêm một loạt các COD khác nhau là beauvericin B, C; allobeauvericin A, B, C. Từ các nghiên cứu này cho thấy, các loài nấm thuộc chi Paecilomyces có tiềm năng lớn trong sản xuất COD [126].
Chỉ duy nhất nấm Hirsutella kobayasii trong chi Hirsutella được công bố về khả năng sinh tổng hợp COD. Khi nuôi cấy Hirsutella kobayasii, Vongvanich và cs đã thu nhận và xác định được một COD mới thuộc nhóm hexadepsipeptide là Hirsutellide A [190]. Cho đến nay chưa có thêm một loài nào thuộc chi Hirsutella này được ghi nhận có khả năng sinh tổng hợp COD.
Ngoài các chi nấm ký sinh côn trùng đáng chú ý ở trên, có nhiều loài thuộc các chi nấm khác cũng có khả năng sinh tổng hợp COD như Halosarphia sp., Pseudoxylaria sp., Lecanicillium sp., Xylaria sp., Aspergillus terreus, Mycelia sterilia, Trichoderma asperellum, Conidiobolus… Bassianolide có thể được phân lập từ Lecanicillium sp. và Xylaria sp. BCC1067 [84]. Một loạt các octadepsipeptide phổ rộng PF1022A, PF1022B, PF1022C, PF1022D, PF1022E, PF1022F, PF1022G, PF1022H đã
được tìm thấy ở chủng Mycelia sterilia sp. PF1022 và Trichoderma asperellum [45, 129]. Hay gần đây nhất sáu hợp chất COD mới là pseudoxylallemycin (A-F) được ghi nhận phân lập khi nuôi cấy chủng nấm
Pseudoxylaria sp. X802 [216]; beauvericin và enniatin A, B được tách chiết từ nấm Conidiobolus coronatus [133]…
1.2.2.2. Nấm Cordyceps spp. sinh tổng hợp cyclooligomer depsipeptide Cordyceps là một chi nấm thuộc họ Cordycipitaceae, bộ Hypocreales,
lớp nấm Ascomycetes, ngành Ascomycota [166]. Cordyceps lần đầu tiên được các nhà khoa học Trung Quốc phát hiện ở vùng núi cao nguyên Tây Tạng. Đây là chi nấm nổi tiếng về sự đa dạng về số lượng loài, phổ ký chủ cũng như các ứng dụng trong y học [40]. Hiện nay có 183 loài Cordyceps đã được ghi nhận [145]. Các loài thuộc chi Cordyceps phân bố chủ yếu ở vùng núi cao ở độ cao 3.600 - 4.000 m so với mực nước biển, được tìm thấy ở Bắc Mỹ, Châu Âu và Châu Á, chủ yếu là ở các nước như Trung Quốc, Nhật Bản, Nepal, Bhutan, Việt Nam, Hàn Quốc và Thái Lan [40].
Có thể bạn quan tâm!
-

 Nghiên cứu đa dạng và sinh tổng hợp Cyclooligomer depsipeptide của nấm ký sinh côn trùng tại Khu Bảo tồn thiên nhiên Copia và Vườn quốc gia Xuân Sơn - 2
Nghiên cứu đa dạng và sinh tổng hợp Cyclooligomer depsipeptide của nấm ký sinh côn trùng tại Khu Bảo tồn thiên nhiên Copia và Vườn quốc gia Xuân Sơn - 2 -
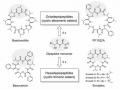 Các Hợp Chất Trao Đổi Thứ Cấp Từ Nấm Ký Sinh Côn Trùng
Các Hợp Chất Trao Đổi Thứ Cấp Từ Nấm Ký Sinh Côn Trùng -
 Khả Năng Ức Chế Sự Sinh Trưởng Của Tế Bào Ung Thư Của Một Số Cod
Khả Năng Ức Chế Sự Sinh Trưởng Của Tế Bào Ung Thư Của Một Số Cod -
 Tách Chiết, Tinh Sạch Và Nghiên Cứu Cấu Trúc Cyclooligomer Depsipeptide Từ Nấm Ký Sinh Côn Trùng
Tách Chiết, Tinh Sạch Và Nghiên Cứu Cấu Trúc Cyclooligomer Depsipeptide Từ Nấm Ký Sinh Côn Trùng -
 Phương Pháp Tuy N Chọn Chủng Nấm Sinh Cyclooligomer Depsipeptide
Phương Pháp Tuy N Chọn Chủng Nấm Sinh Cyclooligomer Depsipeptide -
 Đánh Giá Hoạt Tính Gây Độc Tế Bào Theo Phương Pháp Mtt
Đánh Giá Hoạt Tính Gây Độc Tế Bào Theo Phương Pháp Mtt
Xem toàn bộ 228 trang tài liệu này.
Thành phần của chất chuyển hóa của Cordyceps làm cho chúng có thể chịu được các điều kiện khắc nghiệt đặc trưng ở độ cao (nhiệt độ thấp, thiếu oxy và tiếp xúc với bức xạ tia cực tím) [130]. Khi nghiên cứu về chi nấm Cordyceps, ở cả các loài tự nhiên và nhân tạo người ta đã phát hiện được rất nhiều chất chuyển hóa thứ cấp đáng chú ý như là nucleosides, sterols flavonoids, cyclic peptides, phenolic, bioxanthracenes, polyketides và alkaloids [130]. Nhưng theo Kuo và cs rất hiếm khi COD được tìm thấy trong chi Cordyceps [90].
Tuy nhiên những năm gần đây, trong nghiên cứu của Wang và cs đã báo cáo rằng loài C. cicadae có khả năng tổng hợp beauvericin E, beauvericin J, beauvericin và beauvericin A. Trong nghiên cứu này, lần đầu tiên cordycepin A, beauvericins E và J được báo cáo từ chi Cordyceps, ghi nhận rằng C. cicadae có thể là một nguồn tổng hợp COD thuộc nhóm hexadepsipeptide mới [193].
Trong một nghiên cứu khác của Rachmawati và cs cũng xác định khả năng sinh beauvericin của loài C. militaris [139]. Hay trong quá trình tìm kiếm các loại thuốc chống lại tế bào ung thư, ba hợp chất COD mới là cardinalisamides A, B, C đã được phân lập từ việc nuôi cấy chủng nấm ký sinh côn trùng C. cardinalis NBRC 103832 [180]. Có thể thấy, chi Cordyceps là một trong những nguồn vi sinh vật sinh tổng hợp COD tiềm năng.
1.2.3. Con đường sinh tổng hợp cyclooligomer depsipeptide ở nấm
Cyclooligomer depsipeptide (COD) là những peptide có phổ hoạt tính sinh học rộng được sản sinh từ các vi sinh vật thông qua con đường sinh tổng hợp peptide ngoài ribosome bởi các enzyme phức hợp (module) chứa một hay nhiều protein lớn (NRPSs) như là enniatin synthetase (ESyn), beauvericin synthetase (BeauvSyn), bassianolide synthetase (BassSyn) và PF1022 synthetase (PFSyn) [160, 169, 170]. Mỗi module bao gồm một miền C (Condensation - Ngưng tụ), một miền A (Adenylation - Kích hoạt acid amin phụ thuộc ATP bằng cách adenyl hóa acid cacboxylic tạo thành aminoacyl adenylate) và miền PCP (Peptidyl carrier protein - protein vận chuyển peptidyl). Ngoài ra module 2 sẽ liên kết với một miền đầu cuối có tên là PCP2b-C3 (xem hình 1.5) [25].
Tại module 1, một D-hydroxycarboxylic acid được kích hoạt tại miền A1 và liên kết cộng hóa trị với miền PCP1. Các L-amino acid được kích hoạt miền A của module thứ 2 (A2), được liên kết với miền PCP2a liền kề và bị methyl hóa tại miền Mt (Methylation - Methyl hóa). Các D- hydroxycarboxylic acid và L-amino acid sau khi được kích hoạt bởi module 1 và module 2 sẽ bắt cặp với nhau cho đến khi tạo thành các hexa- hoặc octapeptidol (gọi là sự kéo dài chuỗi hình thành các depsipeptide) [25].
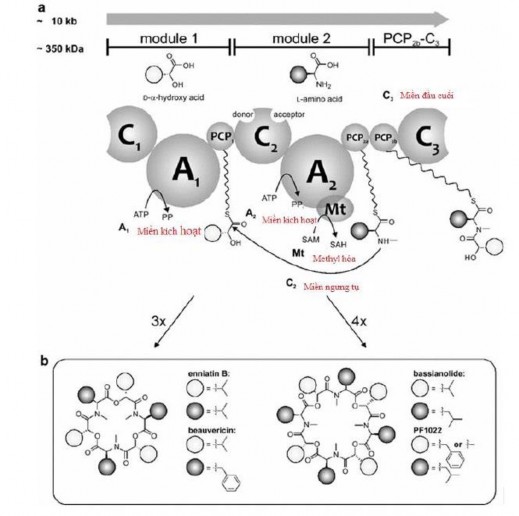
Hình 1.5. Sinh tổng hợp COD [25]
Có hai mô hình khác nhau đã được đề xuất về sự kéo dài chuỗi hình thành các depsipeptide. Thứ nhất là mô hình „„parallel‟‟. Trong mô hình này, chuỗi depsipeptide hình thành bằng cách thêm một dipeptidol bao gồm một hydroxycarboxylic acid và một amino acid đã bị methyl hóa đã được ghép nối với nhau trong miền C2. Miền PCP2a hoặc miền PCP2b hoạt động như một “vị trí chờ” cho đến khi dipeptidol tiếp theo được hình thành. Sau đó, sự hình thành liên kết este và cấu trúc vòng sẽ được diễn ra ở miền C3 [25]. Thứ hai là mô hình “linear” hoặc “looping”. Trong mô hình này, quá trình ngưng tụ của D-hydroxycarboxylic acid vẫn diễn ra tại miền PCP1. Sau đó sự kéo dài của các dipeptidol được chuyển qua các miền PCP1 và PCP 2a/b. Sự hình
thành liên kết peptide được xúc tác trong miền C2 còn miền C3 xúc tác cho sự hình thành liên kết este và tạo nên cấu trúc vòng. Tuy nhiên trong mô hình này, vai trò của hai miền PCP2a/b vẫn chưa rõ ràng, vì chỉ cần một trong hai miền đã đủ để sinh tổng hợp sản phẩm cuối cùng. Đã có giả thuyết cho rằng miền C1 không có chức năng xúc tác trực tiếp vì khi các enzyme bị cắt ngắn thiếu miền C1 vẫn có thể hoạt động bình thường. Do đó, miền C1 có thể đóng vai trò trong việc ổn định hoặc hỗ trợ quá trình xúc tác [164, 209].
Hiện tại, không rõ mô hình nào trong hai mô hình trên thể hiện chính xác hoạt động của NRPSs và cần phải có các nghiên cứu thêm để có thể hiểu đầy đủ về cơ chế cơ bản của quá trình sinh tổng hợp COD ở nấm.
1.3. Nhu cầu dinh dưỡng và điều kiện nuôi cấy của nấm ký sinh côn trùng sinh tổng hợp cyclooligomer depsipeptide
Khả năng sinh tổng hợp các loại COD từ nấm phụ thuộc nhiều vào điều kiện dinh dưỡng và nuôi cấy. Do đó, nghiên cứu nhằm tối ưu hóa môi trường dinh dưỡng và điều kiện nuôi cấy là vô cùng cần thiết trong sản xuất các chất chuyển hóa thứ cấp nói chung và các COD nói riêng [194]. Việc lựa chọn môi trường nuôi cấy, cũng như các thay đổi về độ pH môi trường, nguồn cacbon, nguồn nitơ được chứng minh nhân tố đóng vai trò quan trọng trong quá trình sinh tổng hợp COD từ nấm [91, 135, 203].
1.3.1. Lựa chọn môi trường nuôi cấy
Thông thường trong các nghiên cứu về các chất chuyển hóa thứ cấp của nấm ký sinh côn trùng, các nhà khoa học thường sử dụng môi trường lên men rắn để thu hệ sợi và thể quả. Phổ biến nhất như các loài Cordyceps militaris, Isaria tenuipes… được nuôi cấy trên môi trường cơ chất rắn, sau thời gian nhất định thu hệ sợi, thể quả để tách chiết các hoạt chất sinh học [1, 29, 99]. Tương tự, khi nghiên cứu về nấm ký sinh côn trùng sinh COD ban đầu các nhà nghiên cứu cũng sử dụng các môi trường rắn để nuôi cấy thu sinh khối
nấm. Nhóm tác giả Park và cs đã nuôi cấy Cordyceps bassiana trên môi trường cơm gạo (1 gạo: 1 nước) trong thời gian 7 tuần để thu sinh khối nấm [132]. Khi nghiên cứu về khả năng sinh tổng hợp beauvericin của nấm Isaria tenuipes, Supothina và cs nuôi cấy các chủng nấm trên môi trường cơm gạo có bổ sung 2% nhộng tằm trong thời gian 21 ngày [168]. Có thể thấy nuôi cấy nấm ký sinh côn trùng trên giá thể rắn sẽ gặp những hạn chế nhất định. Không phải chủng nấm ký sinh côn trùng nào cũng sinh quả thể khi lên men rắn, việc thu hệ sợi trên môi trường rắn gặp khó khăn hơn nhiều so với trên môi trường lỏng. Tiếp đến, chưa tính đến hàm lượng COD tích lũy được trong sinh khối và thể quả thì thời gian nuôi cấy dài trong lên men rắn là một bất lợi. Cũng trong nghiên cứu của mình, tác giả Supothina và cs đã nuôi cấy 4 chủng Isaria tenuipes trong cả môi trường rắn và lỏng nhằm so sánh hàm lượng COD và hiệu suất tổng giữa hai phương pháp. Kết quả cho thấy, ở cả 4 chủng hàm lượng COD tích lũy được và hiệu suất COD tổng trong môi trường lỏng đều cao hơn lên men rắn nhiều lần [168]. Điều này cho thấy, lên men lỏng trong sản xuất COD ở các chủng nấm ký sinh côn trùng là một hướng đi tiềm năng.
Trong các nghiên cứu nuôi cấy các chủng nấm ký sinh côn trùng sinh tổng hợp COD trong môi trường lên men lỏng đã công bố, các nhà khoa học sử dụng rất đa dạng các loại môi trường khác nhau. Thông thường nấm nói chung và nấm ký sinh côn trùng nói riêng thường được nuôi cấy trong môi trường Sabouraud (SBR) hay môi trường dịch chiết khoai tây (PDA, PDB). Nhưng trong khảo sát môi trường phù hợp cho sinh tổng hợp COD của các chủng nấm khác nhau các nhà nghiên cứu sử dụng rất nhiều các loại môi trường khác nhau. Khi tối ưu hóa môi trường cho sự sinh trưởng và sinh tổng hợp beauvericin cho chủng Fusarium oxysporum KFCC 11363P, Lee và cs đã sử dụng 10 loại môi trường khác nhau bao gồm: PDB, MB, FCM, FNM, YMB, NS, FBM, CzD, FDM, MCzD [91]. Hay gần đây khi nghiên cứu một
số COD từ nấm Conidiobolus coronatus, Paszkiewicz và cs đã lựa chọn môi trường cho chủng nghiên cứu từ môi trường MM, LB và các môi trường MM, LB cải tiến [133]. Các kết quả trong những nghiên cứu trên cũng cho thấy, khả năng sinh trưởng và sinh tổng COD của cùng một chủng nấm nghiên cứu trong các môi trường dinh dưỡng khác nhau rất khác nhau. Điều này chứng minh tầm quan trọng của lựa chọn môi trường nuôi cấy cho sự sinh trưởng và sinh tổng hợp COD của các chủng nấm ký sinh côn trùng.
1.3.2. Độ pH
Độ pH của môi trường nuôi cấy là một trong những yếu tố quan trọng nhất bởi vì nó có ảnh hưởng rõ rệt đến sự phân ly của các ion, cấu trúc và hoạt động của một số enzyme xúc tác phản ứng trao đổi chất, cũng như gây ảnh hưởng đáng kể đến các hiện tượng sinh lý phức tạp như tính thấm của màng và hình thái tế bào. Sự thay đổi độ pH ban đầu ảnh hưởng đến nhiều quá trình tế bào như điều hoà và sinh tổng hợp sinh các chất chuyển hóa thứ cấp của vi sinh vật nói chung và của nấm nói riêng [31, 144]. Chính vì vậy, độ pH môi trường nuôi cấy giữ vai trò quyết định đến quá trình sinh trưởng, phát triển và sinh tổng hợp COD từ nấm.
Trong các tài liệu đã công bố, khả năng sinh trưởng và sinh tổng hợp COD của mỗi loài nấm ký sinh côn trùng khác nhau lại thích nghi với môi trường có độ pH rất khác nhau. Điển hình như C. coronatus sinh trưởng và sinh một số loại COD hiệu quả nhất trong môi trường có độ pH bằng 5 [133]. Chủng nấm ký sinh côn trùng Aschersonia sp. lại sinh trưởng tốt nhất ở độ pH 7 [159]. Trong khi dải pH tối ưu cho sự sinh tổng hợp beauvericin của các loài thuộc chi Fusarium hoặc chi Beauveria là 7,0 -7,2 [56, 135].
Ngay trong các chủng khác nhau trong cùng một loài, độ pH thích hợp cho sự sinh trưởng cũng rất khác nhau. Dải pH tối ưu cho các chủng Cordyceps militaris trong các nghiên cứu của Rózsa và cs [146] hay Adnan
và cs [14] là 4-5,5. Trong khi chủng Cordyceps militaris trong Geng và cs lại thích nghi với môi trường pH bằng 7,5 [58].
Chính vì vậy, việc nghiên cứu ảnh hưởng của độ pH môi trường đối với sự sinh trưởng cũng như khả năng sinh tổng hợp COD của các chủng nấm ký sinh côn trùng là rất cần thiết.
1.3.3. Nguồn cacbon
Khả năng đồng hóa cacbon giữa các chủng nấm ký sinh côn trùng là không giống nhau, ngoài ra cần căn cứ vào mục đích của quá trình nuôi cấy để lựa chọn nguồn cacbon phù hợp. Quá trình sinh tổng hợp COD có thể được thực hiện theo nhiều con đường khác nhau và dưới sự tác động trực tiếp hoặc gián tiếp của nhiều loại enzyme liên quan. Tuy nhiên việc sử dụng con đường sinh tổng hợp COD nào lại do nguồn cacbon có mặt trong môi trường tại thời điểm đó chi phối.
Đơn cử như trong quá trình sinh tổng hợp beauvericin (Hình 1.6), có thể thấy mối liên hệ chặt chẽ giữa nguồn cacbon và con đường sinh tổng hợp beauvericin. Theo Xu và cs, có bảy nguồn cacbon tiềm năng cho quá trình sinh tổng hợp beauvericin bao gồm glucose, sucrose, fructose, maltose, xylose, lactose và arabinose. Trong đó glucose là nguồn cacbon hiệu quả nhất cho quá trình sinh tổng hợp beauvericin của chủng nấm nghiên cứu [203].
Các kết quả nghiên cứu gần đây đối với nấm Beauveria bassiana cho thấy các nguồn cacbon dễ chuyển hóa, đặc biệt là glucose rất thích hợp cho sự sinh trưởng cũng như sản xuất các chất chuyển hóa thứ cấp của loài này [16, 148]. Bên cạnh các hexose và ribose, các chủng nấm ký sinh côn trùng cũng có thể sử dụng các nguồn cacbon phức tạp hơn như tinh bột từ khoai tây, sắn, khoai lang, ngô, gạo [136]. Tuy nhiên, ngoài các nghiên cứu sử dụng môi trường có chứa dịch chiết khoai tây thì hiện chưa có nghiên cứu nào về sự sinh tổng hợp COD của các nấm ký sinh côn trùng trên các nguồn cacbon kể trên.