oxy hóa [207,208]. Trong nghiên cứu này, EPS giàu phosphate có thể được sản xuất bởi L. plantarum VAL6 trong điều kiện stress pH 3. Thực tế khi phân tích khả năng chống oxy hóa thông qua hoạt động bắt gốc tự do DPPH của EPS được sản xuất bởi
L. plantarum VAL6 dưới các điều kiện stress môi trường, ngoại trừ EPS được sản xuất dưới điều kiện stress nhiệt ở 42 oC, các EPS khác đều thể hiện hoạt động bắt gốc tự do cao hơn so với EPS được sản xuất ở điều kiện nuôi cấy bình thường. Ngoài ra, kết quả cũng cho thấy EPS được sản xuất dưới điều kiện stress pH 3 cho hoạt tính chống oxy hóa cao nhất (14 µmol TE/g) (Hình 3.19).
a
b
c
c
c
d
e
Bắt gốc tự do DPPH (µmol TE/g)
16
14
12
10
8
6
4
2
0
EPSn EPS42 EPS47 EPSpH3 EPSpH8 EPSNaCl EPSCO2
Loại EPS
Hình 3.19. Khả năng chống oxy hóa của các loại EPS được sản xuất bởi L. plantarum VAL6 dưới các điều kiện stress môi trường. EPSn (không gây stress); EPS42 (stress nhiệt ở 42 oC); EPS47 (stress nhiệt ở 47 oC), EPSpH3 (stress ở pH 3), EPSpH8 (stress ở pH 8), EPSNaCl (stress NaCl), EPSCO2 (tăng nồng độ CO2). Các chữ cái khác nhau trên đầu cột thể hiện sự khác biệt có ý nghĩa thống kê với p<0,05
Ngoài nhóm phosphate thì hàm lượng protein trong EPS cũng chịu trách nhiệm một phần cho hoạt tính chống oxy hoá của nó [209]. Hoạt tính chống oxy hoá của protein là do các thành phần acid amin như tyrosine, methionine, histidine, vv…, những acid amin này cho proton tới các gốc tự do [210]. Kết quả của hiện tại cho thấy hàm lượng protein trong EPS được phân lập từ L. plantarum VAL6 dưới các điều kiện stress môi trường cao hơn đáng kể so với đối chứng. Đặc biệt, hàm lượng protein trong EPS cao nhất (6,71 mg/g EPS) khi L. plantarum VAL6 được stress ở pH 3 (Hình 3.20), tương ứng với hoạt tính chống oxy hóa của EPS cao nhất (Hình 3.19).
Một điều khá thú vị được phát hiện trong nghiên cứu này là stress CO2 lại kích thích làm tăng mật số L. plantarum VAL6 (Bảng 3.2). Trong khi đó, mật số của L. plantarum VAL6 khác biệt không có ý nghĩa thống kê (p>0,05) dưới các điều kiện stress ở 42 oC, 47 oC, pH 8, và NaCl so với trường hợp không gây stress. Ngoài ra, mặt dù EPS được tổng hợp với hàm lượng cao nhất nhưng mật số của L. plantarum VAL6 trong điều kiện stress ở pH 3 lại thấp nhất (6,24 Log CFU/mL), nhưng kết quả này chứng minh L. plantarum VAL6 có thể sống sót trong điều kiện pH 3. Một số nghiên cứu trước đây cũng chứng minh L. plantarum có thể tồn tại trong điều kiện pH thấp. Lee và cs. (2011) báo cáo rằng một số chủng L. plantarum được phân lập từ kim chi Hàn Quốc có thể tồn tại ở pH 2,5 trong 2 giờ [211]. Tương tự, mật số của L. plantarum ZDY 2013 thay đổi không đáng kể ở pH 3 và nó có thể tồn tại trong 6 giờ ở pH 2 với mật số trên 7 Log CFU/mL [212].
a
b
dc
c
c
d
e
Hàm lượng protein (mg/g EPS)
8
7
6
5
4
3
2
1
0
EPSn EPS42 EPS47 EPSpH3 EPSpH8 EPSNaCl EPSCO2
Loại EPS
Hình 3.20. Hàm lượng protein trong EPS được sản xuất bởi L. plantarum VAL6 dưới các điều kiện stress môi trường. EPSn (không gây stress); EPS42 (stress nhiệt ở 42 oC); EPS47 (stress nhiệt ở 47 oC), EPSpH3 (stress ở pH 3), EPSpH8 (stress ở pH 8), EPSNaCl (stress NaCl), EPSCO2 (tăng nồng độ CO2). Các chữ cái khác nhau trên đầu cột thể hiện sự khác biệt có ý nghĩa thống kê với p<0,05
Khi so sánh khả năng sống sót sau sấy đông khô, kết quả nghiên cứu đã tìm thấy có sự tương quan tỷ lệ thuận giữa lượng EPS tổng hợp được và khả năng sống sót sau sấy đông khô (Bảng 3.2). Với năng suất EPS đạt cao nhất, stress ở pH 3 cũng cho tỷ lệ sống sót sau sấy đông khô cao nhất (hơn 30%). Tỷ lệ sống sót sau đông khô lần lượt đạt 25,19 và 22,79% tương ứng với điều kiện nuôi cấy tăng
cường CO2 và stress ở pH 8. Trong trường hợp stress NaCl, ở 42 và 47 oC tỷ lệ này dao động trong khoảng 6-10%. Một điểm đáng chú ý là tỷ lệ sống sót sau sấy đông khô của vi khuẩnL. plantarum VAL6 không được gây stress chỉ đạt 0,01% mặt dù hàm lượng EPS đo được là 8 g/L. Như đã mô tả, EPS là một trong những thành phần tham gia vào cấu tạo vách tế bào của vi khuẩn Lactobacillus. Do đó ở điều kiện không gây stress vẫn có thể đo được một lượng EPS nhất định. Tuy nhiên, lượng EPS này chưa đủ để tạo thành lớp vi bao bảo vệ vi khuẩn dưới tác động của quá trình sấy đông khô. Điều này giải thích tại sao khả năng sống sót sau sấy đông khô của L. plantarum VAL6 không được gây stress là rất thấp. Đối với vi khuẩn L. plantarum VAL6 được gây stress, sự kích thích bởi stress môi trường tạo thành lớp EPS xung quanh tế bào, đó như một màng bảo vệ giúp vi khuẩn tránh bị mất nước và sự oxy hoá màng trong quá trình sấy. Nhiều nghiên cứu trước đây cũng đã chứng minh dưới tác động của stress môi trường tế bào vi khuẩn sẽ phản ứng bằng cách tạo ra các hợp chất bảo vệ, chính các hợp chất này sẽ giúp cho vi khuẩn tăng tỷ lệ sống sót trong quá trình sấy và bảo quản sinh khối [213].
Ngoài hoạt tính chống oxy hoá, đặc tính prebiotic của EPS cũng nhận được nhiều sự quan tâm. EPS có thể thúc đẩy sự phát triển của vi khuẩn probiotic và duy trì cân bằng hệ vi sinh đường ruột. Tiềm năng prebiotic của EPS được sản xuất bởi LAB cũng đã được xác nhận [214,215] bởi vì nó được sử dụng bởi các vi khuẩn probiotic [216]. Do đó, nghiên cứu đã đánh giá tiềm năng prebiotic của EPS được sản xuất bởi L. plantarum VAL6 dưới các điều kiện stress môi trường khác nhau dựa trên sự phát triển của các vi khuẩn probiotic như L. acidophilus, L. plantarum và B. longum trên môi trường MRS không đường có bổ sung các loại EPS này. Kết quả chứng minh EPS được sản xuất bởi L. plantarum VAL6 dưới các điều kiện stress và không gây stress đều có khả năng kích thích sự phát triển của vi khuẩn probiotic (Bảng 3.3).
Theo kết quả bảng 3.3, khi sử dụng EPS được sản xuất bởi L. plantarum VAL6 (cả trong điều kiện stress và không gây stress) làm nguồn carbon thay thế, mật số sau 24 giờ nuôi cấy của các vi khuẩn probiotic kiểm tra đều đạt trên 8 Log CFU/mL, so với mật số khi chủng giống ban đầu chỉ khoảng 106 CFU/mL. Không có nhiều khác biệt về mật số vi khuẩn probiotic giữa môi trường MRS thông thường và môi trường MRS không đường có bổ sung EPS. Ngoài ra, khi so sánh khả năng
kích thích probiotic giữa các loại EPS khác nhau, kết quả cho thấy EPSCO2 thể hiện khả năng prebiotic vượt trội hơn so với các loại EPS còn lại (Bảng 3.3). Cũng giống với nghiên cứu này, α -D-glucan (một polysacchride) được tổng hợp bởi L. plantarum DM5 cũng có thể kích thích sự phát triển của vi khuẩn probiotic. Bên cạnh đó, các α -D-glucan này được tiêu hóa thấp bởi dịch dạ dày nhân tạo và có khả năng trì hoãn sự phát triển của vi khuẩn Enterobacteriaceae [217]. Tiềm năng prebiotic của EPS từ Weissiella cibaria, Weissiella confusa, L. plantarum và Pediococcus pentosaceus được tìm thấy khi chúng thể hiện khả năng chống lại sự tiêu hóa của trong dạ dày và ruột, cũng như tăng cường tính chọn lọc của vi khuẩn có lợi, đặc biệt là Bifidobacteria [218].
Bảng 3.3. Mật số vi khuẩn probiotic được nuôi cấy trong môi trường MRS không đường có bổ sung EPS của L. plantarum VAL6
Mật số của vi khuẩn probiotic (Log CFU/mL) | |||
L. acidophilus | L. plantarum | B. longum | |
Đối chứng | 8,14 0,02ab | 8,13 0,02ab | 8,13 0,02c |
EPSn | 8,10 0,04a | 8,17 0,06b | 8,08 0,01ab |
EPS42 | 8,13 0,01ab | 8,17 0,05b | 8,06 0,02a |
EPS47 | 8,19 0,11b | 8,08 0,04a | 8,15 0,03c |
EPSpH3 | 8,15 0,01ab | 8,13 0,01a | 8,12 0,01bc |
EPSpH8 | 8,06 0,02a | 8,14 0,02ab | 8,04 0,03a |
EPSNaCl | 8,07 0,03a | 8,10 0,01a | 8,05 0,04a |
EPSCO2 | 8,06 0,03a | 8,24 0,05c | 8,26 0,01d |
Có thể bạn quan tâm!
-

 Các Phương Pháp Phân Lập Và Tuyển Chọn Vi Khuẩn L. Platarum
Các Phương Pháp Phân Lập Và Tuyển Chọn Vi Khuẩn L. Platarum -
 Phân Lập Và Định Danh Chủng Vi Khuẩn Lactobacillus
Phân Lập Và Định Danh Chủng Vi Khuẩn Lactobacillus -
 Sự Thay Đổi Mật Số L. Plantarum Val6 Dưới Điều Kiện Stress Ph Cùng Với Kiểm Tra Mật Số, Sự Sống Sót Sau Sấy Đông Khô Của L. Plantarum
Sự Thay Đổi Mật Số L. Plantarum Val6 Dưới Điều Kiện Stress Ph Cùng Với Kiểm Tra Mật Số, Sự Sống Sót Sau Sấy Đông Khô Của L. Plantarum -
 Tỷ Lệ Các Monosaccharide: (A) Mannose; (B) Glucose; (C) Galactose; (D) Arabinose; (E) Rhamnose Và (F) Xylose Trong Eps Được Sản Xuất Bởi L. Plantarum Val6 Dưới Điều
Tỷ Lệ Các Monosaccharide: (A) Mannose; (B) Glucose; (C) Galactose; (D) Arabinose; (E) Rhamnose Và (F) Xylose Trong Eps Được Sản Xuất Bởi L. Plantarum Val6 Dưới Điều -
 Điện Di Trên Gel Agarose Sản Phẩm Pcr Mrna Của Các Gen Liên Quan Tổng Hợp Eps Dưới Điều Kiện Stress Ph
Điện Di Trên Gel Agarose Sản Phẩm Pcr Mrna Của Các Gen Liên Quan Tổng Hợp Eps Dưới Điều Kiện Stress Ph -
 Thay Đổi Biểu Hiện Mrna Của Các Gen Liên Quan Tổng Hợp Eps Dưới Tác Động Của Các Thách Thức Môi Trường So Với Điều Kiện Không Gây Stress
Thay Đổi Biểu Hiện Mrna Của Các Gen Liên Quan Tổng Hợp Eps Dưới Tác Động Của Các Thách Thức Môi Trường So Với Điều Kiện Không Gây Stress
Xem toàn bộ 169 trang tài liệu này.
*EPSn (không gây stress); EPS42 (stress nhiệt ở 42 oC); EPS47 (stress nhiệt ở 47 oC), EPSpH3 (stress ở pH 3), EPSpH8 (stress ở pH 8), EPSNaCl (stress ở NaCl), EPSCO2 (tăng nồng độ CO2). Đối chứng là môi trường MRS bình thường không bổ sung EPS. Các chữ cái khác nhau trong cùng một cột thể hiện sự khác biệt có ý nghĩa thống kê với p<0,05.
Trong nghiên cứu này, dựa trên dữ liệu về sự phát triển ổn định của các chủng vi khuẩn probiotic thử nghiệm khi sử dụng EPS do L. plantarum VAL6 sản xuất như nguồn carbon thay thế, kết quả cho thấy các EPS này có thể được khai thác như một prebiotic tiềm năng.
3.3. Ảnh hưởng của stress môi trường lên thành phần monosaccharide của EPS được sản xuất bởi L. plantarum VAL6
3.3.1. Stress nhiệt
Những thay đổi trong điều kiện môi trường được chứng minh làm thay đổi sản xuất polysaccharide ngoại bào. Hơn nữa, mối tương quan giữa thành phần monosaccharide trong EPS và khả năng thích ứng với stress môi trường ở LAB cũng đã được báo cáo [11]. Để rõ hơn về mối tương quan này, nghiên cứu đã phân tích thành phần monosaccharide của EPS được sản xuất bởi L. plantarum VAL6 dưới tác động của stress nhiệt độ (42 và 47 oC). Kết quả cho thấy có sự thay đổi trong thành phần monosaccharide của EPS dưới tác động của stress nhiệt độ cao (Hình 3.21).
Thành phần monosaccharide được ghi nhận bao gồm các loại đường khác nhau chứng minh EPS được sản xuất bởi L. plantarum VAL6 là HePS. Đặc biệt, EPS được sản xuất dưới điều kiện stress nhiệt có tỷ lệ mannose thấp hơn so với điều kiện nuôi cấy thông thường. Cụ thể, trong điều kiện stress ở 42 °C, tỷ lệ mannose dao động trong khoảng 69 đến 80%, trong khi tỷ lệ này tăng đều đặn từ 56,72% sau 1 giờ xử lý lên 73,25% sau 7 giờ xử lý dưới điều kiện stress ở 47 °C, so với khoảng 83% trong điều kiện không gây stress (Hình 3.21A). Kết quả cho thấy nhiệt độ cao làm giảm có ý nghĩa (p<0,05) hàm lượng mannose trong EPS được sản xuất bởi L. plantarum VAL6 (Bảng 3.1.1 phụ lục trang 131).
Ngược lại, tỷ lệ glucose cao hơn được tìm thấy trong thành phần EPS dưới điều kiện stress nhiệt (Hình 3.21B). Đối với stress ở 42 °C, tỷ lệ glucose đạt cao nhất 22,35% sau 5 giờ sau xử lý, và sau đó giảm xuống 16% sau 7 giờ xử lý, nhưng vẫn cao hơn so với đối chứng (khoảng 14%). Đối với stress ở 47 °C, hàm lượng glucose tăng dần từ 17,73% sau 1 giờ xử lý lên 23,92% sau 5 giờ xử lý và sau đó lại giảm. Như vậy kết quả đã chứng minh stress nhiệt làm tăng đáng kể hàm lượng glucose trong thành phần mopnosaccharide của EPS (Bảng 3.1.2 phụ lục trang 131).
Tương tự như với glucose, tỷ lệ galactose sau khi stress nhiệt ở cả nghiệm thức 42 và 47 oC cao hơn so với ở điều kiện không gây stress (Hình 3.21C). Tuy nhiên, tỷ lệ galactose khác nhau không đáng kể (p>0,05, Bảng 3.1.3 phụ lục trang
131) giữa các nhiệt độ xử lý 42 và 47 oC và dao động khoảng 2,5-3,4%, so với
khoảng 1,2% ở nhóm đối chứng không gây stress. Đáng chú ý, sau 5 giờ xử lý ở 42
°C, hàm lượng galactose trong EPS đạt mức cao nhất với 6,58% (Hình 3.21C).
(B)
Không stress 42 °C 47 °C
(A)25
Không stress 42 °C 47 °C
Tỷ lệ mannose (%)
Tỷ lệ glucose (%)
100
80 20
60 15
40 10
20 5
0
0 1 3 5 7
Thời gian (giờ)
0
0 1 3 5 7
Thời gian (giờ)
(C)
Không stress 42 °C 47 °C
Tỷ lệ galactose (%)
7
6
5
4
3
2
1
0
0 1 3 5 7
(E)
10
Không stress 42 °C 47 °C
8
6
4
2
0
0
1
3
5
7
Thời gian (giờ)
Thời gian (giờ)
(D)
Không stress 42 °C 47 °C
Tỷ lệ arabinose (%)
10
8
6
4
2
0
(F)
0 1 3 5 7
Thời gian (giờ)
Không stress 42 °C 47 °C
Tỷ lệ rhamnose (%)
Tỷ lệ xylose (%)
8
7
6
5
4
3
2
1
0
0 1 3 5 7
Thời gian (giờ)
Hình 3.21. Tỷ lệ các monosaccharide: (A) mannose; (B) glucose; (C) galactose; (D) arabinose; (E) rhamnose và (F) xylose trong EPS được sản xuất bởi L. plantarum VAL6 dưới điều kiện stress nhiệt
Tỷ lệ xylose trong EPS cũng bị ảnh hưởng bởi stress nhiệt. Kết quả cho thấy dưới tác động của stress nhiệt, tỷ lệ xylose cao hơn so với điều kiện không gây stress (Hình 3.21F). Theo đó, hàm lượng xylose đạt cao nhất là 6 và 6,55% tương ứng với xử lý ở nhiệt độ 42 và 47 °C khoảng 1 giờ (Hình 3.21F).
Điều thú vị là kết quả ghi nhận sự hiện điện của arabinose trong thành phần EPS dưới điều kiện stress nhiệt, trong khi đường này không được phát hiện trong điều kiện nuôi cấy bình thường (Hình 3.21D). Tỷ lệ arabinose lần lượt là 2,1 và 8,96% tương ứng với xử lý ở 42 và 47 oC trong 1 giờ (Hình 3.21D). Hàm lượng arabinose giảm dần trong suốt thời gian thử nghiệm và không còn phát hiện được sau 7 giờ xử lý ở 42 oC. Tuy nhiên, sau 7 giờ xử lý ở 47 oC, hàm lượng arabinose tăng trở lại (Hình 3.21D). Các kết quả này cho thấy stress nhiệt độ cao có thể kích hoạt L. plantarum VAL6 tổng hợp arabinose và cũng có thể là các loại đường không xác định khác trong thành phần EPS.
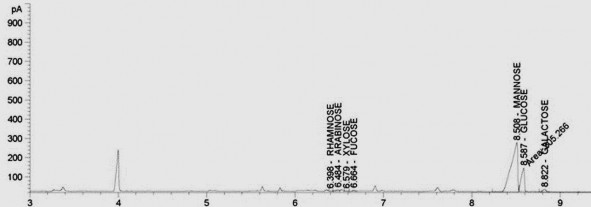
Hình 3.22. Sắc ký đồ thành phần monosaccharide trong EPS được sản xuất bởi L. plantarum VAL6 sau 3 giờ gây stress ở 42 oC
Một kết quả đặc biệt nữa là các loại đường hiếm như rhamnose và fucose được tìm thấy với tỷ lệ cao hơn trong thành phần EPS dưới một số điều kiện xử lý nhất định (Hình 3.21 và Hình 3.22). Cụ thể, xử lý ở 47 oC làm tăng đáng kể tỷ lệ rhamnose (Hình 3.21E). Theo đó, hàm lượng rhamnose đạt tối đa 8% sau 1 giờ xử lý ở 47 oC, cao gấp 10,4 lần so với đối chứng (chỉ 0,7%), và sau đó giảm dần (Hình 3.21E). Đáng chú ý hơn, xử lý ở 42 °C không có bất kỳ sự cải thiện nào về hàm lượng rhamnose trong thành phần EPS, trên thực tế nó thấp hơn so với trong điều kiện nuôi cấy bình thường (Hình 3.21E) nhưng khác biệt này không có ý nghĩa thống kê (p>0,05, Bảng 3.1.4 phụ lục trang 131). Điều này cho thấy có một cơ chế điều chỉnh cụ thể trong con đường sinh tổng hợp EPS được điều khiển chặt chẽ bởi
các kích thích nhiệt. Cuối cùng, đường fucose được hiển thị với một lượng nhỏ khi
L. plantarum VAL6 được gây stress ở 42 °C trong 3 giờ (khoảng 0,47%) và 5 giờ (khoảng 0,25%) (Hình 3.22), trong khi nó không được phát hiện ở các điều kiện xử lý khác. Tổng hợp lại, những kết quả này đã chứng minh thành phần EPS được tổng hợp bởi L. plantarum VAL6 có thể được điều khiển bằng cách kiểm soát nhiệt độ nuôi cấy.
3.3.2. Stress pH
Điều kiện môi trường nuôi cấy có thể làm thay đổi thành phần và tính chất chức năng của EPS tổng hợp được [219,220]. Cùng với nhiệt độ, pH cũng là một trong những yếu tố ảnh hưởng đến tổng hợp EPS ở LAB [221]. Để làm rõ hơn vấn đề này, nghiên cứu đã khảo sát ảnh hưởng của các điều kiện stress ở pH 3 và pH 8 lên thành phần monosaccharide của EPS. Kết quả cho thấy tỷ lệ các đường trong EPS thay đổi đáng kể (Các bảng 3.2.1, 3.2.2 và 3.2.3 phụ lục trang 132) khi so sánh giữa các xử lý gây stress và không gây stress. Tương tự như stress nhiệt, stress pH cũng làm giảm tỷ lệ mannose (Hình 3.23A). Dưới điều kiện stress ở pH 3, tỷ lệ mannose là 83,46% sau 1 giờ xử lý nhưng sau đó giảm xuống 75-76% khi kéo dài thời gian xử lý. Đối với stress ở pH 8, tỷ lệ mannose trong EPS dao động trong khoảng 73-74%, so với 83,44% ở điều kiện không gây stress.
Ngược lại, xử lý stress pH làm tăng hàm lượng glucose (Hình 3.23B). Dưới điều kiện stress ở pH 3, hàm lượng glucose trong EPS tăng từ 12,55% sau 1 giờ xử lý lên tỷ lệ tối đa là 15,77% sau 3 giờ và sau đó giảm dần xuống 14,21% sau 7 giờ xử lý. Trong khi dưới điều kiện stress ở pH 8, tỷ lệ glucose dao động trong khoảng 17,15-18,59%, so với khoảng 14% trong điều kiện không gây stress.
Tỷ lệ galactose trong EPS được sản xuất bởi L. plantarum VAL6 khi tiếp xúc với stress pH 8 tăng mạnh (Hình 3.23C). Dưới tác động của stress pH 8, tỷ lệ galactose dao động từ 6,20-6,49% so với 1,21% của đối chứng. Trái lại, tỷ lệ galactose chỉ khoảng 0,62-1,75% trong điều kiện stress ở pH 3.
Kết quả nghiên cứu cũng tìm thấy sự khác biệt trong hàm lượng rhamnose của EPS khi L. plantarum VAL6 tiếp xúc với các điều kiện stress pH (Hình 3.23E). Đặc biệt, hàm lượng rhamnose trong EPS tăng mạnh ở điều kiện stress pH 3 và tỷ lệ thuận với thời gian gây stress. Tỷ lệ rhamnose sau 7 giờ gây stress đạt xấp xỉ 5%,