Tất cả các chủng phân lập tiết siderophores trong môi trường CAS và khả năng sản xuất siderophore phụ thuộc vào tiềm năng đối kháng của mỗi chủng. Sản xuất siderophore lớn nhất được ghi nhận ở chủng Pseudomonas 6ba6, 6ba2, 6ba3 và 6t14. Trong số 11 chủng đã phân lập có 4 chủng thể hiện hoạt tính protease mạnh (6ba6, 6ba2, 6L10 và 6L14), và 5 chủng có hoạt tính protease yếu (6ba3, 3ss9, ke, 3k9 và mbj) 2 chủng (6t14 và 3tg8) không có hoạt tính protease. Không có chủng nào có khả năng tổng hợp enzyme chitinase và cellulase.
* Xác định thành phần hóa học trong dịch nuôi cấy của chủng vi khuẩn vùng rễ khả năng kháng Phytophthora bằng phân tích LC-MS và GC-MS
Kết quả phân tích GC - MS của chủng RB.DS29 (Bacillus velezensis) được thể hiện qua bảng 3.24 cho thấy, các hợp chất có khả năng bay hơi do chủng Bacillus velezensis RB.DS29 tiết ra trong dịch nuôi cấy có bổ sung bào tử Phytophthora bao gồm: Silane, diethyldodecyloxy (3-phen-); Disulfide, methyl 1-(methylthio); Propanoic acid, 2-methyl-, decyl..; 2-Oxoacetic acid, ethyl ester, o; Ethanol, 2-(2- butoxyethoxy). Tuy nhiên, các hợp chất do chủng Bacillus velezensis RB.DS29 tổng hợp không có trong nghiên cứu Hernández-León và cộng sự (2005). Nghiên của đã phân lập được các chủng Pseudomonas fluorescens UM16, UM240, UM256 và UM270 có khả năng kháng nấm Botritis cinerea nhờ tổng hợp được các hợp chất dễ bay hơi [171]. Kết quả phân tích GC - MS có mặt các chất chứa S như Methanethiol, Dimethyl sulfide, Methyl thiolacetate, ,4-Dithiapentane, S, S-Dimethyl dithiocarbonate, Methyl methylthiomethyl disulfide, Thiazole.
Bảng 3.24. Các hợp chất thứ cấp kháng Phytophthora của của chủng RB.DS29 bằng phân tích GC-MS
Tên hợp chất | RT (phút) | Vùng | Vùng (%) | |
1 | Silane, diethyldodecyloxy (3-phen | 1,034 | 1020196 | 0,55 |
2 | Disulfide, methyl 1-(methylthio) | 1,312 | 77522777 | 41,96 |
3 | Propanoic acid, 2-methyl-, decyl | 1,351 | 31148258 | 16,86 |
4 | 2-Oxoacetic acid, ethyl ester, o | 1,431 | 74779259 | 40,47 |
5 | Ethanol, 2-(2-butoxyethoxy) | 1,708 | 302305 | 0,16 |
Có thể bạn quan tâm!
-

 Ảnh Hưởng Của Chủng Vi Khuẩn Đến Một Số Chỉ Tiêu Sinh Trưởng Và Kháng Bệnh Của Cây Hồ Tiêu Sau 120 Ngày Lây Nhiễm Nấm Fusarium
Ảnh Hưởng Của Chủng Vi Khuẩn Đến Một Số Chỉ Tiêu Sinh Trưởng Và Kháng Bệnh Của Cây Hồ Tiêu Sau 120 Ngày Lây Nhiễm Nấm Fusarium -
 Định Danh Các Chủng Có Tiềm Năng Kháng Phytophthora, Fusarium, Kháng Tuyến Trùng Meloidogyne Gây Bệnh Rễ Ở Cây Hồ Tiêu
Định Danh Các Chủng Có Tiềm Năng Kháng Phytophthora, Fusarium, Kháng Tuyến Trùng Meloidogyne Gây Bệnh Rễ Ở Cây Hồ Tiêu -
 Ảnh Hưởng Của Ph Đến Sinh Trưởng Của Chủng Rb.ds29, Rb.cj41 Và Rb.ek7
Ảnh Hưởng Của Ph Đến Sinh Trưởng Của Chủng Rb.ds29, Rb.cj41 Và Rb.ek7 -
 Mối Tương Quan Giữa Hoạt Tính Enzyme Và Tỷ Lệ Tử Vong Tuyến Trùng
Mối Tương Quan Giữa Hoạt Tính Enzyme Và Tỷ Lệ Tử Vong Tuyến Trùng -
![Ảnh Hưởng Của Hexahydropyrrolo [1,2-A] Pyrazine-1,4-Dione Đến Tỷ Lệ Tử Vong Của Tuyến Trùng Và Tỷ Lệ Nở Của Trứng Tuyến Trùng](https://tailieuthamkhao.com/uploads/2022/12/28/tuyen-chon-nghien-cuu-dac-tinh-khang-tac-nhan-gay-benh-va-tao-che-pham-18-120x90.jpg) Ảnh Hưởng Của Hexahydropyrrolo [1,2-A] Pyrazine-1,4-Dione Đến Tỷ Lệ Tử Vong Của Tuyến Trùng Và Tỷ Lệ Nở Của Trứng Tuyến Trùng
Ảnh Hưởng Của Hexahydropyrrolo [1,2-A] Pyrazine-1,4-Dione Đến Tỷ Lệ Tử Vong Của Tuyến Trùng Và Tỷ Lệ Nở Của Trứng Tuyến Trùng -
 Ảnh Hưởng Của Chất Mang Đến Hoạt Tính Kháng Tuyến Trùng Trong In Vitro Của Chế Phẩm Chứa Chủng Rb.ek7
Ảnh Hưởng Của Chất Mang Đến Hoạt Tính Kháng Tuyến Trùng Trong In Vitro Của Chế Phẩm Chứa Chủng Rb.ek7
Xem toàn bộ 224 trang tài liệu này.
Kết quả phân tích LC-MS (Bảng 3.25) cho thấy, các hợp chất không có khả năng bay hơi do chủng RB.DS29 tiết ra trong dịch nuôi cấy có bổ sung bào tử
Phytophthora bao gồm Metronidazole-oh, Sulfadiazin. Kết quả này tương đồng với nghiên cứu của Bibi và cộng sự (2018) đã phân lập được 6 chủng vi khuẩn có khả năng kháng Phytophthora capsici và Pythium ultimum thông qua cơ chế tiết một số hợp chất kháng nấm. Kết quả phân tích trên hệ thống LC-MS có sự hiện diện của Sulfamethoxypyridazine, Sulfamonomethoxine, Sulfamerazine, Dimetridazole, Sulfadiazin, Metronidazole-oh, Nalidixic acid.
Bảng 3.25. Phân tích LC- MS các hợp chất kháng Phytophthora của chủng RB.DS29
Tên chất | Công thức | RT | Khối phổ | Vùng | |
1 | Metronidazole-oh | C6H9N3O4 | 4.1 | 188.0686 | 3739.4 |
2 | Sulfadiazin | C10H10N4O2S | 4.3 | 250.1196 | 26293.1 |
3.2.2.2. Xác định đặc tính kháng nấm Fusarium của chủng vi khuẩn tuyển chọn
* Ảnh hưởng của dịch nuôi cấy vi khuẩn có bổ sung casein, chitin, β-glucan và bào tử nấm Fusarium sp. đến khả năng ức chế nấm Fusarium sp.
Theo Schoffelmeer và cộng sự (1999), thành tế bào nấm Fusarium được cấu tạo bởi chitin, α-1,3-glucan và β-1,3-glucan ngoài ra còn có protein [173]. Bên cạnh đó, ở nhiều loài vi sinh vật, hoạt tính đối kháng có thể thực hiện thông qua khả năng tổng hợp các hợp chất khác nhau như bacteriocin, enzyme, chất độc, hợp chất bay hơi và các chất khác [174]. Chủng Bacillus subtilis RB.CJ41 được nuôi cấy trong môi trường có bổ sung các cơ chất casein 1%, chitin 1% và β- glucan 0,1% cũng như bào tử nấm Fusarium nhằm cảm ứng tổng hợp enzyme và các hợp chất thứ cấp.
Kết quả bảng 3.26 cho thấy, nếu bổ sung chất cảm ứng nào thì hoạt tính enzyme phân giải chất cảm ứng đó được tổng hợp và chỉ có một loại enzyme tương ứng với chất cảm ứng. Riêng nghiệm thức bổ sung bào tử nấm Fusarium thì vi khuẩn tổng hợp cả ba enzyme là protease, chitinase và β-glucanase. Tổ hợp ba enzyme này cùng tấn công phân hủy sợi nấm, làm sợi nấm đứt gãy thành đoạn, hoặc bị teo lại (hình 3.17A). Trong khi đó, Chaurasia và cộng sự (2005) đã tuyển chọn được chủng B. subtilis (NRRL B-30408) phân lập từ đất vùng rễ của cây chè có khả năng tổng hợp được các hợp chất dễ khuếch tán và dễ bay hơi tác động lên hình thái, cấu trúc hệ sợi và bào tử của nấm Fusarium oxysporum như làm ly giải sợi nấm, hình thành cấu trúc hạt trong sợi nấm, bào tử sưng to và có thành dày [174].
Figueroa-López và cộng sự (2016) đã tuyển chọn được chủng Bacillus subtilis
B13 có hoạt tính enzyme chitinase, glucanase, chủng Bacillus megaterium B22 có
hoạt tính enzyme glucanase và chitinase, chủng Bacillus cereus B24 và B25 có cả ba hoạt tính enzyme chitinase, glucanase và protease [175]. Đây đều là những chủng tiềm năng được phân lập từ cây ngô có khả năng kháng nấm Fusarium verticillioides. Bảng 3.26. Khả năng ức chế nấm bệnh của các dịch nuôi cấy chủng RB.CJ41
Hoạt tính enzyme protease (UI/ml) | Hoạt tính enzyme chitinase (UI/ml) | Hoạt tính enzyme β- glucanase (UI/ml) | HSĐK (%) dịch nuôi cấy không bất hoạt | HSĐK (%) dịch nuôi cấy bất hoạt | |
MT + casein | 0,36±0,03 | - | - | 18,3a | 4,2a |
MT+ chitin | - | 3,96±0,08 | - | 18,3a | 0,0b |
MT + β-glucan | - | - | 0,41±0,04 | 20,8a | 5,0b |
MT + bào tử nấm Fusarium | 0,15±0,01 | 1,32±0,05 | 0,29±0,02 | 20,8a | 6,7a |
MT | - | - | - | 3,3b | 0,0b |
CV% | - | - | - | 9,0 | 30,6 |
P | - | - | - | ** | ** |
Ghi chú: HSĐK (%):hiệu suất đối kháng; MT: môi trường; Các số trung bình trong cùng một cột có chữ cái khác nhau thì khác biệt có ý nghĩa thống kê ở mức p<0,01.
Trong điều kiện không bất hoạt dịch nuôi cấy vi khuẩn, không có sự khác biệt có ý nghĩa thống kê về hiệu suất đối kháng nấm bệnh ở các nghiệm thức có bổ sung chất cảm ứng. Hiệu suất đối kháng nấm bệnh đạt từ 18,3% -20,8%. Tuy nhiên, trong điều kiện bất hoạt dịch nuôi cấy vi khuẩn, có sự khác biệt có ý nghĩa thống kê (p<0,01) về hiệu suất đối kháng giữa các nghiệm thức có bổ sung chất cảm ứng. Hiệu suất đối kháng nấm bệnh giảm còn 0% - 6,7%, đặc biệt, nghiệm thức môi trường có bổ sung chitin hiệu suất đối kháng chỉ còn 0%. Hơn nữa, hiệu suất đối kháng ở nghiệm thức môi trường có bổ sung bào tử nấm bệnh giảm từ 20,8% còn 6,7%. Điều này chứng tỏ, hoạt tính kháng nấm Fusarium của chủng Bacillus subtilis RB.CJ41 có thể do enzyme và các hợp chất thứ cấp tác động lên.
Kết quả trình bày ở hình 3.16 cũng cho thấy, có mối tương quan thuận và chặt giữa hoạt tính enzyme chitinase, protease và β-glucanase và hoạt tính kháng nấm Fusarium với hệ số tương quan lần lượt là R=0,515, R=0,680 và R=0,640 (P<0,05).
Kết quả nghiên cứu của Rathore và cộng sự (2020) cũng tương đồng với kết quả trên [176]. Kết quả nghiên cứu chứng minh rằng có mối tương quan thuận giữa phần trăm ức chế sinh trưởng nấm Fusarium oxysporum f. sp. cumini và hoạt tính enzyme chitinase, protease, β-1,3 glucanase, với hệ số tương quan lần lượt đạt 0,991, 0,703 và 0,644. Bên cạnh đó, nghiên cứu cũng chỉ ra có mối tương quan chặt giữa các enzyme với nhau cụ thể là protease – chitinase, protease và β-1,3 glucanase, chitinase và β-1,3 glucanase với hệ số tương quan lần lượt đạt R=0,768, R=0,987, R=0,725 (P=0,01). Sự kết hợp của các enzyme này đã ức chế mạnh mẽ lên sự sinh trưởng của nấm bệnh.
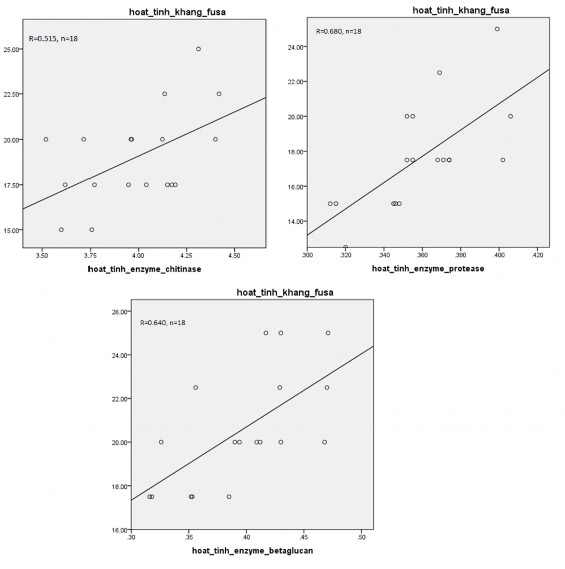
Hình 3.16. Mối tương quan giữa hoạt tính enzyme và hoạt tính kháng Fusarium
Theo Chang và cộng sự (2010), chitinase xúc tác quá trình thủy phân liên kết 1,4-β D-glycosidic nối các tiểu đơn vị Nacetylglucosaminine của chitin [177]. Chitinase được phân loại thành ba nhóm: endochitinase và hai nhóm exochitinase bao gồm chitobiosidases và β N-acetylhexosaminidase. Endochitinase phân cắt chitin một
cách ngẫu nhiên, tạo ra các đa phân tử GlcNAc hòa tan có trọng lượng phân tử thấp, chẳng hạn như chitobiose, chitotriose và chitotetraose; các chitobiosidase phân cắt chitin để giải phóng từng chất dimer GlcNAc (chitobiose) từ đầu không khử của chuỗi chitin; và β-N-acetylhexosaminidases thủy phân chitobiose, chitotriose và chitotetraose từ đầu không nhân tạo của N-acetylchitooligosaccharides, dẫn đến giải phóng N-acetylglucosamine. Chitinase được tinh sạch từ chủng B. subtilis NPU 001 có trọng lượng phân tử 31kDa với hoạt tính enzyme đạt 1,25U/mg và có hoạt tính kháng nấm F. oxysporum. Bên cạnh đó, Khan và cộng sự (2018) đã tuyển chọn được chủng B. subtilis 30VD-1 có khả năng kháng nấm Fusarium sp. và sinh trưởng trên môi trường có chứa chitin nhờ hoạt tính enzyme chitinase, tạo vòng sáng halo xung quanh khuẩn lạc của chúng [11]. Nghiên cứu cũng xác định được các hợp chất kháng nấm Fusarium sp. gồm có phenol, 1-decanol, 1-dodecanol và benzaldehyde.
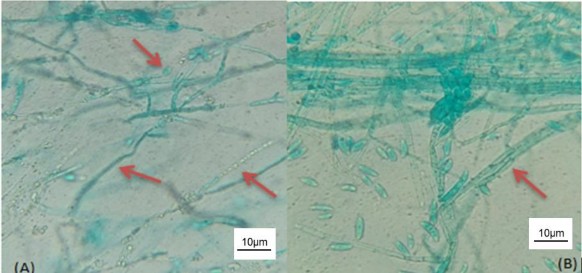
Hình 3.17. Hệ sợi nấm Fusarium dưới tác động của dịch nuôi cấy vi khuẩn RB.CJ41khi bổ sung bào tử nấm Fusarium sp.
(A) Sợi nấm tại đĩa đối kháng (B) sợi nấm tại đĩa đối chứng
* Xác định thành phần hóa học trong dịch nuôi cấy của chủngvi khuẩn vùng rễ tuyển chọn có khả năng kháng nấm Fusarium bằng phân tích LC-MS và GC-MS
Chủng RB.CJ41 được lựa chọn để xác định hợp chất có khả năng kháng nấm Fusarium bằng phương pháp GC-MS và LC-MS. Đây là chủng kháng nấm mạnh nhất khi tuyển chọn các chủng kháng nấm Fusarium mạnh trong in vitro và trong điều kiện vườn ươm. Dịch nuôi cấy vi khuẩn có bổ sung bào tử nấm bệnh có hiệu suất đối kháng với nấm bệnh cao.
Kết quả phân tích GC - MS của chủng Bacillus subtilis RB.CJ41 được thể hiện
qua bảng 3.27, các hợp chất có khả năng bay hơi do chủng Bacillus subtilis RB.CJ41 tiết ra trong dịch nuôi cấy có bổ sung bào tử nấm Fusarium sp. bao gồm 39 chất. Tuy nhiên, các chất có peak rõ nhất là những hợp chất Cyclotrisiloxane, hexamethyl; Pentasiloxane, dodecamethyl; Cyclohexasiloxane, dodecamethyl; 3-Isopropoxy- 1,1,1,7,7,7-hexamethyl; 1 3,4-Dihydroxyphenylglycol, 4TMS; Cyclononasiloxane, octadecamethyl; Cyclotrisiloxane, hexamethyl; Octasiloxane. Kết quả của nghiên cứu này không chứa các hợp chất có trong nghiên cứu của Niku‐Paavola và cộng sự (1999) khi phân tích GC-MS từ dịch nuôi vi khuẩn Lactobacillus plantarum VTT E
‐78076 thu được các chất bao gồm benzoic acid (CAS 65-85-0), 5‐methyl‐2,4‐ imidazolidinedione (CAS 616-03-5, methylhydantoin), tetrahydro‐4‐hydroxy‐4‐ methyl‐2H‐pyran‐2‐one (CAS 674-26-0, mevalonolactone) and 3‐(2‐methylpropyl)‐ 2,5‐piperazinedione (CAS 5845-67-0, cyclo(glycyl‐L ‐leucyl). Các hợp chất cũng hoạt tính kháng nấm Fusarium avenaceum (Gibberella avenacea) VTT D ‐ 80147 [178]. Wang và cộng sự (2012) đã phân lập được chủng vi khuẩn Lactobacillus plantarum IMAU10014 tạo ra phổ rộng các hợp chất kháng nấm, tất cả đều có hoạt tính kháng nấm gây bệnh trên thực vật. Các hợp chất kháng nấm chính gồm 3- phenyllactic acid, Benzeneacetic acid, 2-propenyl ester được thực hiện bằng phân tích HPLC, LC-MS, GC-MS, NMR [179].
Bảng 3.27. Kết quả phân tích GC-MS của chủng Bacillus subtilis RB.CJ41 chứa 8 chất có các đỉnh (peak) rõ
Tên hợp chất | RT (phút) | Vùng | Vùng (%) | |
1 | Cyclotrisiloxane, hexamethyl | 5,076 | 159018 | 0,78 |
2 | Pentasiloxane, dodecamethyl | 9,547 | 256828 | 1,27 |
3 | Cyclohexasiloxane, dodecamethyl | 15,022 | 1767969 | 8,72 |
4 | 3-Isopropoxy-1,1,1,7,7,7-hexamethyl | 17,284 | 4382913 | 21,61 |
5 | 1 3,4-Dihydroxyphenylglycol, 4TMS | 19,273 | 1257123 | 6,20 |
6 | Cyclononasiloxane, octadecamethyl | 20,990 | 387528 | 1,91 |
7 | Cyclotrisiloxane, hexamethyl | 21,168 | 178770 | 0,88 |
8 | Octasiloxane | 22,498 | 153650 | 0,76 |
Tương tự, Sheoran và cộng sự (2015) đã phân lập được chủng Pseudomonas putida BP25 từ rễ cây hồ tiêu có khả năng kháng Phytophthora capsici và tuyến trùng Radopholus similis thông qua cơ chế tiết một số hợp chất kháng nấm và tuyến trùng [180]. Kết quả phân tích trên hệ thống GC-MS có sự hiện diện của một số hợp chất
thơm 1-Undecene; Disulfide dimethyl; Pyrazine, methyl-Pyrazine, 2,5-dimethyl-; Rượu Isoamyl; Pyrazin, metyl-; Dimethyl trisulfide. Ngoài ra chủng Pseudomonas putida BP25 còn có khả năng kháng nấm Magnaporthe oryzae thông qua việc tổng hợp các hợp chất dễ bay hơi pyrazine là 2-methylpyrazine và 2-ethyl-3,6-dimethylpyrazine; cả hai đều ức chế tất cả các giai đoạn phát triển như sự nảy mầm của bào tử, sự phát triển hệ sợi và sự hình thành bào tử của M. oryzae. Để xác nhận hoạt tính kháng nấm của 2- methylpyrazine và 2-ethyl-3,6-dimethylpyrazine đối với bệnh đạo ôn trên cây lúa, trong các thí nghiệm trên cây lúa đã được tiến hành trên hạt lúa và cây con. Tỷ lệ bệnh đạo ôn và mức độ nghiêm trọng của bệnh trên cây con được xử lý bằng hợp chất bay hơi đã giảm đáng kể so với đối chứng. Pyrazine được sử dụng để khử trùng hạt lúa nhờ hoạt tính kháng nấm cũng như khả năng bảo vệ cây con nhằm đảm bảo cây con khỏe mạnh và sạch bệnh [181].
Phân tích LC-MS của chủng Bacillus subtilis RB.CJ41
Kết quả phân tích LC - MS cho thấy, các hợp chất không có khả năng bay hơi do chủng RB.CJ41 tiết ra trong dịch nuôi cấy có bổ sung bào tử nấm Fusarium bao gồm Nalicdic Acid và Metronidazole. Bibi và cộng sự (2018) đã phân lập được 6 chủng vi khuẩn có khả năng kháng Phytophthora capsici và Pythium ultimum thông qua cơ chế tiết một số hợp chất kháng nấm [172]. Kết quả phân tích trên hệ thống LC-MS có sự hiện diện của Sulfamethoxypyridazine, Sulfamonomethoxine, Sulfamerazine, Dimetridazole, Sulfadiazin, Metronidazole-oh, Nalidixic acid. Venturini và cộng sự (2011) chứng minh các hợp chất amphotericin B, itraconazole, voriconazole kết hợp với azithromycin, ciprofloxacin, fluvastatin, ibuprofen, metronidazole có khả năng kháng lại 23 chủng Fusarium sp.[182].
Bảng 3.28. Kết quả phân tích LC-MS của chủng Bacillus subtilis RB.CJ41
Tên chất | Công thức | RT | Khối phổ | Vùng | |
1 | Metronidazole-oh | C6H9N3O4 | 4.0 | 188.0679 | 3635.5 |
2 | Nalidixic acid | C12H12N2O3 | 4.1 | 233.0953 | 13518.3 |
Như vậy, khi phân tích LC-MS từ dịch nuôi cấy chủng Bacillus subtilis
RB.CJ41 xuất hiện 2 chất có khả năng kháng nấm Metronidazole và Nalidixic acid.
3.2.2.3. Xác định đặc tính kháng tuyến trùng Meloidogyne sp. của chủng vi khuẩn ngoại sinh vùng rễ tuyển chọn
* Ảnh hưởng của dịch nuôi cấy vi khuẩn có bổ sung casein, chitin, và tuyến trùng đến
tỷ lệ tử vong và tỷ lệ nở của trứng tuyến trùng Meloidogyne sp. theo thời gian
Protease là enzyme có khả năng phá vỡ các liên kết peptide trong protein và được chia thành hai loại dựa trên vị trí cắt của các liên kết peptide: endopeptidase và exopeptidase. Endopeptidase xúc tác sự phân hủy các liên kết peptide trong chuỗi polypeptide, trong khi exopeptidase là nguyên nhân phá vỡ liên kết peptide ở cuối chuỗi polypeptide. Enzyme protease ngoại bào được sản xuất bởi vi khuẩn đóng vai trò quan trọng trong việc kiểm soát một số loại mầm bệnh thực vật trong đó có tuyến trùng [183-185]. Bên cạnh protease, vi khuẩn vùng rễ còn có khả năng sản sinh chitinase để kháng lại nấm bệnh, tuyến trùng. Enzyme thủy phân liên kết β-1,4 giữa N-acetyl glucosamine (NacGlc) trên chitin là polysacharide của thành tế bào ở nấm và tuyến trùng [186]. Protease và chitinase có khả năng làm thoái hóa lớp biểu bì của tuyến trùng và đóng vai trò như chất diệt tuyến trùng nhằm kiểm soát sinh học quần thể tuyến trùng [187].
Bảng 3.29. Ảnh hưởng của dịch nuôi cấy chủng RB.EK7 đến tỷ lệ tử vong (%) tuyến trùng Meloidogyne sp.
Hoạt tính enzyme (UI/ml) 12 giờ 24 giờ 48 giờ
Nghiệm thức | Chitinase | Protease |
MT+ tuyến trùng | 1,79±0,07 | 0,089±0,001 |
MT+ casein | - | 0,20±0,02 |
MT+chitin | 4,5±0,10 | - |
ĐC nước | - | - |
ĐC môi trường | - | - |
P | - | - |
CV % | - | - |
KBH | BH | KBH | BH | KBH | BH |
83,67 | 78,33 | 88,33 | 93,67 | 95,67 | 100,0 |
±1,41a | ±12,82a | ±2,11a | ±5,95a | ±4,31a | ±0,000a |
71,00 | 82,67 | 82,00 | 92,00 | 100,0 | 97,67 |
±4,17a | ±6,61a | ±5,51a | ±1,73a | ±0,000a | ±2,07a |
79,333 | 68,33 | 81,00 | 78,00 | 93,67 | 96,67 |
±8,98a | ±6,47a | ±3,80a | ±4,83b | ±5,53a | ±5,77a |
4,33 | 6,33 | 7,67 | 7,33 | 8,00 | 8,00 |
±1,32b | ±0,87b | ±3,15c | ±1,34d | ±1,42c | ±2,47c |
12,00 | 23,00 | 29,67 | 22,67 | 23,67 | 29,33 |
±4,48ab | ±1,80b | ±5,19b | ±0,53c | ±1,61b | ±3,57b |
** | ** | ** | ** | ** | ** |
16,90 | 14,70 | 7,20 | 5,19 | 4,45 | 5,55 |
Ghi chú: MT: môi trường, KBH (Không bất hoạt enzyme); BH (Bất hoạt enzyme), ĐC: đối chứng. ** Khác biệt có ý nghĩa thống kê ở mức p<0,01, các chữ cái giống nhau trên cùng một cột thể hiện sự khác biệt không có ý nghĩa thống kê theo trắc nghiệm phân hạng Duncan’s Multiple Rang Test.
Kết quả bảng 3.29 cho thấy, dịch nuôi cấy chủng RB.EK7 có bổ sung tuyến trùng có hoạt tính enzyme tương đối thấp (hoạt tính chitinase và protease đạt lần lượt 1,79 UI/ml, 0,089 UI/ml) so với dịch nuôi cấy có bổ sung cơ chất casein (hoạt tính