và RB.CJ41, 6 chủng vi khuẩn vùng rễ có hoạt tính kháng tuyến trùng Meloidogyne bao gồm RB.EK7, RB.CJ4, RB.EK2, RB.DS33, RB.BH11, RB.CS30 trong điều kiện in vitro. Các chủng này được sử dụng cho nghiên cứu định danh các chủng có tiềm năng kháng Phytophthora, Fusarium và tuyến trùng Meloidogyne gây bệnh rễ ở cây hồ tiêu.
3.1.3. Định danh các chủng có tiềm năng kháng Phytophthora, Fusarium, kháng tuyến trùng Meloidogyne gây bệnh rễ ở cây hồ tiêu
Các chủng vi khuẩn vùng rễ của cây hồ tiêu bao gồm RB.DS29, RB.CJ41 và RB.EK7 có hoạt tính kháng mạnh nhất đối với Phytophthora, Fusarium và tuyến trùng Meloidogyne trong điều kiện vườn ươm được lựa chọn để định danh theo khóa phân loại của Bergey (Bảng 3.15).
Bảng 3.15. Đặc điểm hình thái và đặc tính sinh hóa của các chủng có hoạt tính kháng nấm và tuyến trùng cao
RB.DS29 | RB.CJ41 | RB. EK7 | |
Hình dạng khuẩn lạc | Tròn, không đều, nhăn nheo | Tròn, mép răng cưa | Tròn, nhăn nheo, có vành ở mép |
Màu sắc khuẩn lạc | Trắng | Trắng | Trắng |
Hình dạng tế bào | Que | Que | Que |
Gram* | + | + | + |
Nội bào tử | + | + | + |
Khả năng di động | + | + | + |
Catalase | + | + | + |
Urease | - | - | - |
Sucrose | + | + | + |
Glucose | + | + | + |
Trehanose | + | + | + |
Malnitose | + | + | + |
Nitrate | - | + | - |
Citrate | - | + | - |
Indol | - | - | - |
VP | + | + | + |
MR | + | - | |
ONPG | + | + | |
Arginine | - | - | - |
Có thể bạn quan tâm!
-

 Hiệu Quả Kháng Phytophthora Capsici Trên Đoạn Thân Hồ Tiêu
Hiệu Quả Kháng Phytophthora Capsici Trên Đoạn Thân Hồ Tiêu -
 Ảnh Hưởng Của Chủng Vi Khuẩn Đến Sinh Trưởng Rễ Của Cây Hồ Tiêu Sau 90 Ngày Lây Nhiễm Phytophthora
Ảnh Hưởng Của Chủng Vi Khuẩn Đến Sinh Trưởng Rễ Của Cây Hồ Tiêu Sau 90 Ngày Lây Nhiễm Phytophthora -
 Ảnh Hưởng Của Chủng Vi Khuẩn Đến Một Số Chỉ Tiêu Sinh Trưởng Và Kháng Bệnh Của Cây Hồ Tiêu Sau 120 Ngày Lây Nhiễm Nấm Fusarium
Ảnh Hưởng Của Chủng Vi Khuẩn Đến Một Số Chỉ Tiêu Sinh Trưởng Và Kháng Bệnh Của Cây Hồ Tiêu Sau 120 Ngày Lây Nhiễm Nấm Fusarium -
 Ảnh Hưởng Của Ph Đến Sinh Trưởng Của Chủng Rb.ds29, Rb.cj41 Và Rb.ek7
Ảnh Hưởng Của Ph Đến Sinh Trưởng Của Chủng Rb.ds29, Rb.cj41 Và Rb.ek7 -
 Các Hợp Chất Thứ Cấp Kháng Phytophthora Của Của Chủng Rb.ds29 Bằng Phân Tích Gc-Ms
Các Hợp Chất Thứ Cấp Kháng Phytophthora Của Của Chủng Rb.ds29 Bằng Phân Tích Gc-Ms -
 Mối Tương Quan Giữa Hoạt Tính Enzyme Và Tỷ Lệ Tử Vong Tuyến Trùng
Mối Tương Quan Giữa Hoạt Tính Enzyme Và Tỷ Lệ Tử Vong Tuyến Trùng
Xem toàn bộ 224 trang tài liệu này.
Ghi chú: +: có, -: không có, * Gram dương (+), Gram (-)
Dựa trên hệ thống phân loại của Bergey [154] cho thấy, cả 3 chủng đều thuộc chi Bacillus với các đặc điểm như tế bào hình que, có khả năng sinh nội bào tử, có khả năng di động, tổng hợp enzyme catalase, lên men một số loại đường. Tuy nhiên, chi Bacillus gồm có 142 loài, trong đó chỉ có 95 loài được định danh dựa vào các phản ứng sinh hóa. Mỗi loài có rất nhiều phản ứng sinh hóa khác nhau nên để xác định chính xác tên loài phải mất nhiều thời gian, công sức và chi phí. Vì vậy, để định danh chính xác tên loài của các chủng vi khuẩn vùng rễ đã tuyển chọn, nghiên cứu tiến hành kết hợp với phương pháp sinh học phân tử bằng cách giải trình tự gen vùng 16S rRNA để đảm bảo độ tin cậy cho kết quả.

Hình 3.10. Hình dạng khuẩn lạc và tế bào của 3 chủng RB.DS29, RB.CJ41 và RB.EK7
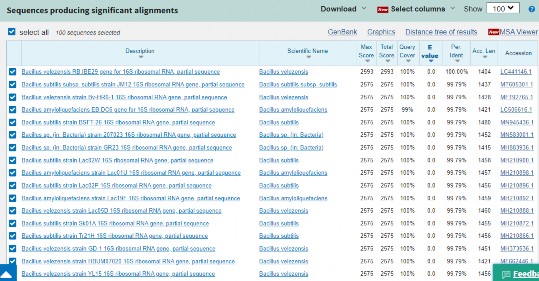
Kết quả giải trình tự gen 16S rRNA của 3 chủng RB.DS29, RB.CJ41 và RB.EK7 và so sánh trình tự gen 16S rRNA bằng chương trình BLASTn search trên NCBI được trình bày tại bảng 3.16.
Bảng 3.16. Kết quả định danh bằng phương pháp giải trình tự gen 16S rRNA các chủng có hoạt tính kháng nấm và tuyến trùng cao
Chủng | Chiều dài trình tự (bp) | Loài | Tương đồng (%) | Accession number | |
1 | RB.DS29 | 1404 | Bacillus velezensis | 100% | LC441166 |
2 | RB.CJ41 | 1397 | Bacillus subtilis | 100% | LC602155 |
3 | RB.EK7 | 1418 | Bacillus velezensis | 100% | LC602156 |
Đối với chủng RB.DS29 (có hoạt tính kháng Phytophthora): Trình tự nucleotide (1404 bp) của gen 16S rRNA của chủng có độ bao phủ và tương đồng là 100% với các trình tự gen 16S rRNA của Bacillus velezensis S141 (AP018402). Kết quả phân tích
phát sinh loài bằng phần mềm MEGA 6.0 (hình 3.14) cho thấy chủng RB.DS29 (RB.IBE29: theo tên gọi Viện Công nghệ Sinh học và Môi trường, IBE) có quan hệ di truyền gần gũi nhất với Bacillus velezensis.
Hình 3.11. Kết quả so sánh mức độ tương đồng của chủng RB.DS29 với các chủng vi khuẩn trên cơ sở dữ liệu NCBI
Đối với chủng RB.CJ41 (có hoạt tính kháng nấm Fusarium): Trình tự nucleotide (1397 bp) của gen 16S rRNA của chủng CJ41 có độ bao phủ và tương đồng là 100% với các trình tự gen 16S rRNA của Bacillus subtilis H1 (CP026662), B. subtilis PZ-24 (MT184836), B. subtilis MK729017.1 (MT111087), B. subtilis MN727117.1 (MT111025), B. subtilis LC520135 (MT111012), B. subtilis QT180-9 (MT083981), B. subtilis QH14-11 (MT078611), B. subtilis DL3 (MT043898), B.
subtilis CpI4 (MN960273). Kết quả phân tích phát sinh loài bằng phần mềm MEGA
6.0 (hình 3.14) cho thấy chủng RB.CJ41 có quan hệ di truyền gần gũi nhất với
Bacillus subtilis.

Hình 3.12. Kết quả so sánh mức độ tương đồng của chủng RB.CJ41 với các chủng vi khuẩn trên cơ sở dữ liệu NCBI
Đối với chủng RB.EK7 (có hoạt tính kháng tuyến trùng): Trình tự nucleotide (1418 bp) của gen 16S rRNA của chủng RB.EK7 có độ bao phủ và tương đồng là 100% với các trình tự gen 16S rRNA của Bacillus velezensis RA-9 (MG371986). Kết quả phân tích phát sinh loài bằng phần mềm MEGA 6.0 (hình 3.14) cho thấy chủng RB.EK7 có quan hệ di truyền gần gũi nhất với B. velezensis.

Hình 3.13. Kết quả so sánh mức độ tương đồng của chủng RB.CJ41 với các chủng vi khuẩn trên cơ sở dữ liệu NCBI
Kết quả định danh cho thấy, cả 3 chủng tuyển chọn có hoạt tính kháng nấm và tuyến trùng đều thuộc chi Bacillus, là một tác nhân sinh học đầy tiềm năng trong phòng trừ bệnh hại cây trồng. Chúng có khả năng hình thành nội bào tử, dễ dàng xâm chiếm rễ cây, sản xuất kháng sinh phổ rộng và thúc đẩy sinh trưởng thực vật, sống xót khi xử lý nhiệt và sấy khô, sản xuất chế phẩm dạng bột với mật độ ổn định và thời gian lâu dài [153]. Các loài thuộc chi Bacillus có khả năng tổng hợp các chất chuyển hóa thứ cấp với hoạt tính sinh học cao ức chế sự phát triển của mầm bệnh thực vật và vi sinh vật gây hại tại vùng rễ [155]. Hơn nữa, Bacillus sp. là chi chiếm chủ yếu trong hệ vi sinh vật vùng rễ thực vật, tác động rất ít đến thành phần của các cộng đồng vi khuẩn ở rễ thực vật [156]. Vì thế, các chủng thuộc chi Bacillus đã được thương mại hóa để sản xuất thuốc trừ sâu sinh học nhằm mục đích kích kiểm soát bệnh trên cây trồng.
Một số loại thuốc trừ sâu sinh học có nguồn gốc từ Bacillus sp. như Kodiak, Compmate, Serenade và Rhapsody hiện đang được sử dụng để kiểm soát nhiều bệnh ở thực vật [157]. Một số loài được phát triển thành các sản phẩm thương mại để kiểm soát bệnh thực vật và tuyến trùng. Chế phẩm BioNem – WP/Biosafe của AgroGreen chứa vi khuẩn B. firmus có khả năng kháng tuyến trùng M. incognita, M. hapla, Heterodera sp., Tylenchulus semipenetrans, Xiphinema index và Ditylenchus dipsaci [158]. Chế phẩm dạng lỏng BioYield của Gustafson bao gồm hai chủng B. velezensis
IN937 và B. subtilis GB03 được dùng để kiểm soát bệnh phát sinh từ đất và giảm mật độ tuyến trùng trên cây cà chua [159]. Chế phẩm Nemix của AgriLife/Chr. Hansen chứa Bacillus sp. được sử dụng để kiểm soát bệnh do tuyến trùng sần rễ trên cây rau và cây ăn trái [142]. VoTiVO được thương mại hóa bởi tập đoàn Bayer CropScience chứa chủng Bacillus firmus GB-126 được dùng để xử lý hạt nhằm kiểm soát tuyến trùng ký sinh thực vật trê cây ngô, cây bông, cây đậu nành, cây cải đường [160]. Pathway Consortia của hãng Pathway Holdings, một sản phẩm chứa vi khuẩn B. subtilis, B. licheniformis, B. megaaterium, B. coagulans, Pseudomonas fluorescens, Streptomyces sp. và Trichoderma, có khả năng kháng tuyến trùng sần rễ [161].

Hình 3.14. Cây phân loại thể hệ mối liên quan giữa chủng RB.DS29, RB.CJ41 và RB.EK7 với các loài gần gũi dựa trên trình tự 16S rRNA
Kết quả nghiên cứu cho thấy chủng vi khuẩn RB.DS29 và RB.EK7 được định danh là Bacillus velezensis là một loại vi khuẩn hiếu khí, gram dương, hình thành nội bào tử, thúc đẩy sinh trưởng ở thực vật cũng nhu khả năng ức chế lại sự phát triển nhiều bệnh ở thực vật do vi khuẩn, nấm và tuyến trùng gây ra [162]. Khi phân tích bộ gen của B. velezensis cho thấy, chúng mang các gen đặc hiệu liên quan đến sinh tổng hợp các chất chuyển hóa thứ cấp, đóng vai trò quan trọng trong cả ức chế mầm bệnh và thúc đẩy tăng trưởng thực vật như gen mã hóa cho quá trình tổng hợp lipopeptide cyclic (tức là, Surfactin, bacillomycin-D, fengycin và bacillibactin) và polyketide (tức
là, macrolactin, bacillaene và difficidin). Mặc dù, đều có tên khoa học là Bacillus velezensis nhưng chủng RB.DS29 lại có khả năng kháng Phytophthora trong khi đó chủng RB.EK7 lại có khả năng kháng tuyến trùng trong điều kiện in vitro và ex vitro. Điều này có thể là do các gen liên quan đến sinh tổng hợp các hợp chất thứ cấp được biểu hiện trong những điều kiện dinh dưỡng và hóa lý của môi trường khác nhau.
Ngoài ra, các chất chuyển hóa thứ cấp được tổng hợp bởi B. velezensis cũng có thể kích hoạt hệ thống kháng cảm ứng ở thực vật. Điều này cũng được khẳng định trong nghiên cứu của Xiang và cộng sự (2017) khi đánh giá tiềm năng của 662 chủng vi khuẩn vùng rễ kích thích tăng trưởng thực vật đối với Meloidogyne incognita J2 trong điều kiện in vitro, nhà lưới, microplot và thử nghiệm trên đồng ruộng [163]. Kết quả chỉ ra rằng, các chủng PGPR gây tỷ lệ tử vong tuyến trùng M. incognita từ 0 đến 100 %, trung bình 39 %. 212 trong số 662 chủng có khả năng gây ra tỷ lệ tử vong
M. incognita J2 cao hơn đáng kể so với nhóm đối chứng, trong đó chi Bacillus có tỷ lệ tử vong cao hơn khi so sánh với các chi khác. Kết quả thử nghiệm trong nhà lưới ghi nhận, chủng B. velezensis Bve2 đã làm giảm trứng tuyến trùng M. incognita/ gram rễ cây bông sau 45 ngày trồng khi so sánh với đối chứng xử lý với chất diệt tuyến trùng Abamectin và Clothianidin kết hợp với B. Firmus I-1582.
Khi tiến hành thử nghiệm trong microplot, Bacillus mojavensis chủng Bmo3,
B. velezensis chủng Bve2, B. subtilis subsp. subilis chủng Bsssu3 và hỗn hợp Abamectin + Bve2 + B. altitudinis chủng Bal13 có khả năng ức chế trứng M. incognita/ g rễ sau 45 ngày trồng. Các chủng Bacillus velezensis Bve2 và Bve12 cũng làm tăng năng suất hạt bông trong thử nghiệm microplot và đồng ruộng. Nhìn chung, kết quả chỉ ra rằng các chủng B. velezensis Bve2 và Bve12, B. mojavensis chủng Bmo3 và hỗn hợp Abamectin + Bve2 + B. altitudinis chủng Bal13 có khả năng làm giảm mật độ quần thể M. incognita và thúc đẩy sinh trưởng của cây bông.
Kết luận nội dung 1:
Từ 269 chủng vi khuẩn đã phân lập tại vùng rễ cây hồ tiêu ở 3 tỉnh Tây Nguyên, nghiên cứu đã tuyển chọn được ba chủng vi khuẩn vùng rễ có khả năng kháng bệnh rễ trên cây hồ tiêu bao gồm chủng Bacillus velezensis RB.DS29 kháng Phytophthora capsici (tác nhân gây bệnh chết nhanh), chủng Bacillus subtilis RB.CJ41 kháng nấm Fusarium oxysporum và chủng Bacillus velezensis RB.EK7 kháng tuyến trùng Meloidogyne sp. (tác nhân gây bệnh chết chậm) trong điều kiện in vitro và ex vitro.
Ba chủng này tiếp tục được nghiên cứu đặc tính kháng nấm và tuyến trùng nhằm làm rõ cơ chế kháng bệnh rễ trên cây hồ tiêu. Đồng thời, cả ba chủng RB.DS29, RB.CJ41, RB.EK7 được dùng làm vật liệu để bước đầu sản xuất chế phẩm phòng trừ bệnh chết nhanh, bệnh chết chậm trên cây hồ tiêu ở Tây Nguyên.
3.2. Nghiên cứu đặc tính và xác định hoạt chất kháng Phytophthora/Fusarium, tuyến trùng Meloidogyne của các chủng vi khuẩn tiềm năng được tuyển chọn
3.2.1. Nghiên cứu điều kiện nhân sinh khối của các chủng vi khuẩn vùng rễ tuyển chọn
3.2.1.1. Môi trường nuôi cấy
Môi trường nuôi cấy là các cơ chất dinh dưỡng được pha chế nhân tạo nhằm đáp ứng cho yêu cầu sinh trưởng, phát triển và sản sinh các sản phẩm trao đổi chất của vi khuẩn. Môi trường dinh dưỡng là yếu tố quan trọng cho sự tăng sinh khối của vi khuẩn [120]. Kết quả khảo sát môi trường thích hợp cho sinh trưởng của các chủng RB.DS29, RB.CJ41 và RB.EK7 được thể hiện ở bảng 3.17 và 3.18.
Đối với chủng RB.DS29: Mật độ vi khuẩn dao động từ 152 x 106 – 1491 x 106 CFU/ml và có sự khác biệt có ý nghĩa thống kê (p<0,01) giữa các nghiệm thức. Nghiệm thức sử dụng môi trường LB có mật độ vi khuẩn cao nhất đạt 1491 x 106CFU/ml gấp 9,8 lần so với môi trường NA có mật độ vi khuẩn thấp nhất đạt 152 x 106CFU/ml. Bên cạnh đó môi trường TSB có mật độ vi khuẩn khá cao đạt 970,67 x 106CFU/ml so với những môi trường còn lại.
Bảng 3.17. Ảnh hưởng của môi trường nuôi cấy đến sinh trưởng của chủng RB.DS29
TSB | NA | LB | Pepton | Pepton 1 | Pepton 2 | Pepton 3 | Pepton 4 | |
Mật độ vi khuẩn (x106CFU/ml) | 970,67 ±69,06b | 152 ±15,87f | 1491,00 ±127,74a | 348 ±15,87cd | 448± 3,46c | 362 ±36,50cd | 180 ±18,33ef | 298 ±13,86d |
Ghi chú: Các số trung bình trong cùng một hàng có chữ cái khác nhau thì khác biệt có ý nghĩa thống kê ở mức p<0,01.
Đối với chủng RB.CJ41: Mật độ vi khuẩn ở các nghiệm thức môi trường khác nhau có sự khác biệt có ý nghĩa thống kê ở mức p<0,01. Môi trường M6 (Glucose 2g/l, Cao thịt 5g/l, Pepton 3g/l, (NH4)2SO4 4g/l, MgSO4.7H2O 0,5g/l) có mật độ vi khuẩn cao nhất (2457 x106 CFU/ml) gấp 5,08 lần mật độ vi khuẩn ở môi trường M4 (Glucose 1g/l, cao nấm men 2,5g/l, cao thịt 2,5g/l, (NH4)2SO4 2g/l, MnCl2.4H2O 0,25g/l, MgSO4.7H2O 0,25g/l) thấp nhất đạt 404,00 x106 CFU/ml. Tuy nhiên, trong nghiên cứu của Posada-Uribe và cộng sự (2015), chủng Bacillus subtilis EA-CB0575 sinh trưởng tốt nhất trong môi trường có thành phần gồm pepton 3g/l, cao thịt 5g/l,
MnCl2.4H2O 0,5g/l, KH2PO4 6g/l, MgSO4.7H2O 0,5g/l, mật độ vi khuẩn đạt 2,01x109CFU/ml [133].
Đối với chủng RB.EK7: Mật độ vi khuẩn dao động từ 160 x 106 – 1313,67 x 106 CFU/ml và có sự khác biệt có ý nghĩa thống kê (p<0,01) giữa các nghiệm thức môi trường. Nghiệm thức M2 (Glucose 2g/l, cao nấm men 5g/l, Pepton 3g/l, KH2PO4 6g/l, (NH4)2SO4 4g/l, MnCl2.4H2O 0,5g/l) có mật độ cao nhất đạt 1313,67 x106 CFU/ml gấp 1,11 -8,21 lần so với các môi trường còn lại.
Bảng 3.18. Ảnh hưởng của môi trường nuôi cấy đến sinh trưởng của chủng RB.CJ41 và RB.EK7
Mật độ vi khuẩn (x106CFU/ml) | ||
Chủng RB.CJ41 | Chủng RB.EK7 | |
M1 | 660,00±33,00c | 1176,00±21,00b |
M2 | 597,67±55,37c | 1313,67±42,77a |
M3 | 1115,33±49,17b | 260,00±22,27d |
M4 | 404,00±143,16d | 189,00±14,00e |
M5 | 1119,00±36,37b | 512,00±36,66c |
M6 | 2457,00±72,38a | 160,00±12,00e |
P | ** | ** |
CV% | 5,23 | 4,14 |
Ghi chú: ** Khác biệt có ý nghĩa thống kê ở mức p<0,01. Các chữ cái giống nhau trên cùng một cột thể hiện sự khác biệt không có ý nghĩa thống kê theo trắc nghiệm phân hạng Duncan’s Multiple Rang Test.
3.2.1.2. pH
pH của môi trường ảnh hưởng đến họat động sống của vi khuẩn do làm thay đổi sự cân bằng về trao đổi chất giữa môi trường và vi khuẩn, có thể giết chết vi khuẩn. Mỗi loại vi khuẩn chỉ thích hợp với một giới hạn pH nhất định, đa số là ở pH trung tính (pH = 7), bởi vì pH nội bào của tế bào sống là trung tính. Kết quả đánh giá ảnh hưởng của pH đến sinh trưởng của các chủng RB.DS29, RB.CJ41 và RB.EK7 được thể hiện ở bảng 3.19.
Đối với chủng RB.DS29: giá trị pH của môi trường biến động từ 5 – 9 dẫn đến biến động mật độ vi khuẩn giao động từ 145,33 x 106 – 3299,33 x 106 CFU/ml và có sự khác biệt có ý nghĩa thống kê giữa các nghiệm thức (p<0,01). Mật độ vi khuẩn tăng dần từ môi trường có pH 5 đến pH 7,0 sau đó lại giảm dần về nghiệm thức pH = 7,5, 8,0, 8,5, 9,0. Ở nghiệm thức môi trường có pH = 7 có mật độ vi khuẩn cao nhất