protease 0,2 UI/ml) và chitin (hoạt tính protease đạt 4,5 UI/ml). Tại thời điểm 12 giờ, môi trường bổ sung tuyến trùng và môi trường bổ sung chitin, tỷ lệ tử vong của tuyến trùng đều giảm khi bất hoạt dịch nuôi cấy. Tuy nhiên, tại thời điểm 24 giờ, chỉ có môi trường bổ sung chitin tỷ lệ tử vong của tuyến trùng giảm 1,12 lần (dịch chưa bất hoạt đạt 81 %, dịch bất hoạt đạt 78 %). Như vậy, enzyme chitinase có thể tác động đến tỷ lệ tử vong của tuyến trùng tại thời điểm này. Tại thời điểm 48 giờ, không có sự tác động của enzyme đến tỷ lệ tử vong của tuyến trùng.
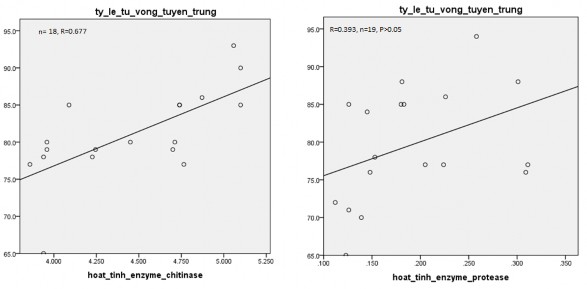
Hình 3.18. Mối tương quan giữa hoạt tính enzyme và tỷ lệ tử vong tuyến trùng
Bên cạnh đó, khi đánh giá mối tương quan giữa hoạt tính enzyme chitinase và protease đến tỷ lệ tử vong (%) tuyến trùng Meloidogyne sp. (hình 3.18) cũng cho thấy, enzyme chitinase và tỷ lệ tử vong tuyến trùng có mối tương quan thuận và rất chặt với hệ số tương quan là R=0,677 (p<0,01). Điều này chứng tỏ, khi hoạt tính enzyme tăng sẽ tăng tỷ lệ tử vong của tuyến trùng. Kết quả của Soliman và cộng sự (2019) ghi nhận có mối tương quan giữa enzyme chitinase đến tỷ lệ tử vong của tuyến trùng [187]. Hoạt tính enzynme chitinase cao nhất ở hai chủng Paenibacillus polymyxa và Bacillus subtilis gây tử vong tuyến trùng Meloidogyne incognita lần lượt đạt 100% và 97%.
Kết quả bảng 3.30 cho thấy, dịch môi trường nuôi cấy vi khuẩn bổ sung tuyến trùng không tác động lên tỷ lệ nở của trứng tại các thời điểm khảo sát. Tại thời điểm 24 giờ, 48 giờ và 72 giờ đều có sự tác động của enzyme chitinase lên tỷ lệ nở của trứng tuyến trùng. Cụ thể trong điều kiện không bất hoạt enzyme, ở nghiệm thức MT+chitin, tỷ lệ nở của trứng tuyến trùng tại thời điểm 24 giờ, 48 giờ và 72 giờ lần lượt đạt 10%, 14%, 17,67%. Hơn nữa, trong điều kiện bất hoạt enzyme, tỷ lệ nở của
trứng tuyến trùng lần lượt đạt 20,67%, 21%, 22,33%, tăng lần lượt 2,06, 1,5, 1,27 lần. Trong khi đó, enzyme protease chỉ tác động vào thời điểm 72 giờ. Ở nghiệm thức MT+casein, ở cả hai điều kiện không bất hoạt và bất hoạt enzyme, tỷ lệ nở của trứng tuyến trùng lần lượt đạt 15,33%, 19,67% tăng 1,28 lần. Như vậy, enzyme chitinase tác động lên tỷ lệ nở của trứng tuyến trùng vào 3 thời điểm 24 giờ, 48 giờ và 72 giờ. Enzyme protease chỉ tác động vào thời điểm 72 giờ.
Bảng 3.30. Ảnh hưởng của dịch nuôi cấy chủng RB.EK7 đến tỷ lệ nở của trứng tuyến trùng Meloidogyne sp.
Hoạt tính enzyme (UI/ml) | 12 giờ | 24h | 48h | 72h | ||||||
Chitinase | Protease | KBH | BH | KBH | BH | KBH | BH | KBH | BH | |
MT+tuyến trùng | 1,79±0,07 | 0,089±0,001 | 13,67 ±4,94b | 0,00 ±0,00c | 19,00 ±1,51b | 6,33 ±1,90c | 20,67 ±1,00b | 10,67 ±3,15b | 25,33 ±1,28b | 12,00 ±1,62b |
MT+casein | - | 0,2±0,02 | 6,67 ±3,06c | 0,00 ±0,00c | 7,33 ±2,73c | 2,33 ±4,28c | 16,00 ±5,58b | 7,33 ±3,98b | 15,33 ±6,70b | 19,67 ±0,44b |
MT+chitin | 4,5±0,10 | - | 2,33 ±2,04c | 0,00 ±0,00c | 10,00 ±2,37bc | 20,67 ±8,08b | 14,00 ±2,31b | 21,00 ±8,59b | 17,67 ±4,36b | 22,33 ±7,73b |
ĐC nước | - | - | 28,67 ±1,71a | 28,67 ±1,71b | 55,33 ±5,03a | 55,33 ±5,03a | 77,67 ±1,66a | 77,67 ±1,66a | 88,67 ±4,07a | 88,67 ±4,07a |
ĐC môi trường | - | - | 32,00 ±1,28a | 32,00 ±1,28a | 52,00 ±4,19a | 52,00 ±4,19a | 78,33 ±5,34a | 78,33 ±5,34a | 86,67 ±4,78a | 86,67 ±4,78a |
P | - | - | ** | ** | ** | ** | ** | ** | ** | ** |
CV % | - | - | 15,02 | 7,45 | 12,16 | 13,17 | 9,27 | 13,62 | 10,11 | 10,24 |
Có thể bạn quan tâm!
-

 Định Danh Các Chủng Có Tiềm Năng Kháng Phytophthora, Fusarium, Kháng Tuyến Trùng Meloidogyne Gây Bệnh Rễ Ở Cây Hồ Tiêu
Định Danh Các Chủng Có Tiềm Năng Kháng Phytophthora, Fusarium, Kháng Tuyến Trùng Meloidogyne Gây Bệnh Rễ Ở Cây Hồ Tiêu -
 Ảnh Hưởng Của Ph Đến Sinh Trưởng Của Chủng Rb.ds29, Rb.cj41 Và Rb.ek7
Ảnh Hưởng Của Ph Đến Sinh Trưởng Của Chủng Rb.ds29, Rb.cj41 Và Rb.ek7 -
 Các Hợp Chất Thứ Cấp Kháng Phytophthora Của Của Chủng Rb.ds29 Bằng Phân Tích Gc-Ms
Các Hợp Chất Thứ Cấp Kháng Phytophthora Của Của Chủng Rb.ds29 Bằng Phân Tích Gc-Ms -
![Ảnh Hưởng Của Hexahydropyrrolo [1,2-A] Pyrazine-1,4-Dione Đến Tỷ Lệ Tử Vong Của Tuyến Trùng Và Tỷ Lệ Nở Của Trứng Tuyến Trùng](https://tailieuthamkhao.com/uploads/2022/12/28/tuyen-chon-nghien-cuu-dac-tinh-khang-tac-nhan-gay-benh-va-tao-che-pham-18-120x90.jpg) Ảnh Hưởng Của Hexahydropyrrolo [1,2-A] Pyrazine-1,4-Dione Đến Tỷ Lệ Tử Vong Của Tuyến Trùng Và Tỷ Lệ Nở Của Trứng Tuyến Trùng
Ảnh Hưởng Của Hexahydropyrrolo [1,2-A] Pyrazine-1,4-Dione Đến Tỷ Lệ Tử Vong Của Tuyến Trùng Và Tỷ Lệ Nở Của Trứng Tuyến Trùng -
 Ảnh Hưởng Của Chất Mang Đến Hoạt Tính Kháng Tuyến Trùng Trong In Vitro Của Chế Phẩm Chứa Chủng Rb.ek7
Ảnh Hưởng Của Chất Mang Đến Hoạt Tính Kháng Tuyến Trùng Trong In Vitro Của Chế Phẩm Chứa Chủng Rb.ek7 -
 Tuyển chọn, nghiên cứu đặc tính kháng tác nhân gây bệnh và tạo chế phẩm phòng trừ bệnh rễ của các chủng vi khuẩn vùng rễ cây hồ tiêu Piper nigrum L. tại Tây Nguyên - 20
Tuyển chọn, nghiên cứu đặc tính kháng tác nhân gây bệnh và tạo chế phẩm phòng trừ bệnh rễ của các chủng vi khuẩn vùng rễ cây hồ tiêu Piper nigrum L. tại Tây Nguyên - 20
Xem toàn bộ 224 trang tài liệu này.
Ghi chú: MT: môi trường, KBH (Không bất hoạt enzyme); BH (bất hoạt enzyme), ĐC: đối chứng. Các chữ cái giống nhau trên cùng một cột thể hiện sự khác biệt không có ý nghĩa thống kê theo trắc nghiệm phân hạng Duncan’s Multiple Rang Test; ** Khác biệt có ý nghĩa thống kê ở mức p<0,01.
Sử dụng vi khuẩn vùng rễ tiết enzyme phân giải như chitinase và protease để ngăn cản quá trình nở của trứng tuyến trùng được nghiên cứu hướng đến nhiều nhất hiện nay. Chitinase có thể phá vỡ sự hình thành lớp chitin ở trứng tuyến trùng và ngăn cản quá trình nở của trứng [187]. Jung và cộng sự (2002) cũng đã chứng minh, vi khuẩn Paenibacillus illinoisensis KJA-424 tấn công vào vỏ trứng tuyến M. incognita sau 48 giờ bằng cách tổng hợp enzyme endo-chitinase làm thay đổi hình dạng của trứng và gây vỡ trứng [188]. Thongkaewyuan và Chairin (2018) ghi nhận enzyme protease từ nấm Metarhizium guizhouense PSUM02 có khả năng ức chế hoàn toàn tỷ lệ nở của tuyến trùng Meloidogyne incognita trong vòng 24 giờ và tỷ lệ tử vong của tuyến trùng trưởng thành sau 48 giờ bằng cách tác động lên lớp vỏ trứng và lớp biểu
bì [189].
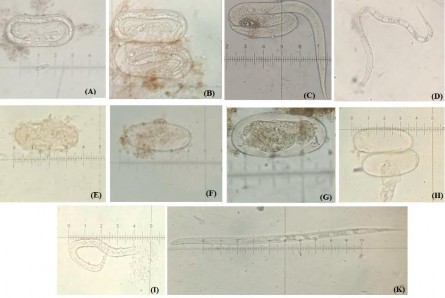
Hình 3.19. Tác động của enzyme ngoại bào của vi khuẩn vùng rễ đến quá trình nở của trứng tuyến trùng và gây chết tuyến trùng trưởng thành
Ghi chú: (A), (B): trứng tuyến trùng bình thường; (C): tuyến trùng nở bình thường; (D) tuyến trùng sống; (E), (F), (G), (H) trứng tuyến trùng bị phá hủy nguyên sinh chất và nhân; (I), (K) tuyến trùng chết.
*Xác định một số nhóm hợp chất có hoạt tính kháng tuyến trùng Meiloidogyne sp. của các chủng vi khuẩn vùng rễ RB.EK7 bằng phân tích LC-MS và GC-MS
Bacillus velezensis có khả năng tổng hợp lipopeptide cyclic (Surfactin, bacillomycin-D, fengycin, và bacillibactin) và polyketide (macrolactin, bacillaene và difficidin). Các chất chuyển hóa thứ cấp được tạo ra bởi Bacillus velezensis cũng có thể gây kích kháng ở thực vật, một quá trình mà thực vật tự bảo vệ mình trước sự xâm nhiễm của các tác nhân lây bệnh. Ngoài ra, Bacillus velezensis còn sản sinh các hợp chất có lợi cho chúng như siderophore, bacteriocins và các hợp chất hữu cơ dễ bay hơi [162].
Kết quả phân tích GC - MS của chủng RB.EK7 (Bacillus velezensis) được thể hiện qua bảng 3.31 cho thấy, các hợp chất có khả năng bay hơi do chủng Bacillus velezensis RB.EK7 tiết ra trong dịch nuôi cấy có bổ sung tuyến trùng bao gồm một số hợp chất chính như Cyclohexasiloxane, dodecamethyl, Pentasiloxane, dodecamethyl-, 2,4-Dihydroxybenzoic acid, 3TMS derivative, 2,4,6- Cycloheptatrien-1-one, 3,5-bis-trimethylsilyl-.
Zhai và cộng sự (2019) đã phân lập được chủng vi khuẩn Pseudomonas putida
có khả năng kháng tuyến trùng M. incognita thông qua cơ chế tiết một số hợp chất
kháng tuyến trùng bao gồm: dimethyl - disulfide, 1 - undecene, 2 –undecene, Nonanone, 2 - octanone, (z) - hexen - 1 - ol axetat, 2 - undecanone, và 1 - (ethenyloxy) Octadecane [190].Tuy nhiên, những hợp chất này không xuất hiện ở dịch nuôi cấy chủng RB.EK7 khi phân tích GC-MS.
Bảng 3.31 Kết quả phân tích GC-MS các hợp chất hoá học do chủng vi khuẩn RB.EK7 tổng hợp
Tên hợp chất | RT (phút) | Vùng | Vùng (%) | |
1 | Cyclohexasiloxane, dodecamethyl | 15.034 | 1860750 | 26,30 |
2 | Pentasiloxane, dodecamethyl- | 17.305 | 1525759 | 21.56 |
3 | 2,4-Dihydroxybenzoic acid, 3TMS derivative | 19.283 | 509194 | 7.20 |
4 | 2,4,6-Cycloheptatrien-1-one, 3,5-bis- trimethylsilyl- | 20.990 | 154118 | 2.18 |
5 | 1,1,1,3,5,5,5-Heptamethyltrisiloxane | 22.613 | 80203 | 1.13 |
6 | 2'-Hydroxy-5'-methylacetophenone, TMS derivative | 26.487 | 136272 | 1.93 |
7 | Silicic acid, diethyl bis (trimethyl bis(trimethylsilyl) ester | 28.905 | 1450378 | 20.50 |
8 | Arsenous acid, tris(trimethylsilyl) ester | 29.188 | 43106 | 0.61 |
9 | 2'-Hydroxy-5'-methylacetophenone, TMS derivative | 29.345 | 141265 | 2.00 |
10 | 2'-Hydroxy-5'-methylacetophenone, TMS derivative | 29.533 | 92278 | 1.30 |
11 | Tetrasiloxane, decamethyl- | 30.141 | 494244 | 6.98 |
12 | Cyclotrisiloxane, hexamethyl- | 30.256 | 263045 | 3.72 |
13 | 2'-Hydroxy-5'-methylacetophenone, TMS derivative | 30.339 | 58324 | 0.82 |
Kết luận chung về đặc tính kháng đặc tính kháng Phytophthora, Fusariumvà tuyến trùng của các chủng RB.DS29, RB.CJ41 và RB.EK7:
Chủng Bacillus velezensis RB.DS29 có cơ chế kháng Phytophthora capsici bằng cách tổng hợp các enzyme phân giải thành tế bào nấm (chitinase, protease và β- glucanase) và các hợp chất thứ cấp ở dạng dễ bay hơi (Silane, diethyldodecyloxy (3- phen-); Disulfide, methyl 1-(methylthio); Propanoic acid, 2-methyl-, decyl..; 2- Oxoacetic acid, ethyl ester, o; Ethanol, 2-(2-butoxyethoxy) và không bay hơi
(Metronidazole-oh, Sulfadiazin).
Chủng Bacillus subtilis RB.CJ41 có cơ chế kháng nấm Fusarium oxysporum bằng cách tổng hợp các enzyme phân giải thành tế bào nấm (chitinase, protease, β- glucanase) và các hợp chất thứ cấp ở dạng dễ bay hơi (Cyclotrisiloxane, hexamethyl; Pentasiloxane, dodecamethyl; Cyclohexasiloxane, dodecamethyl; 3-Isopropoxy- 1,1,1,7,7,7-hexamethyl; 1 3,4-Dihydroxyphenylglycol, 4TMS; Cyclononasiloxane, octadecamethyl; Cyclotrisiloxane, hexamethyl; Octasiloxane) và không bay hơi (Metronidazole và Nalidixic acid).
Chủng Bacillus velezensis RB.EK7 có cơ chế kháng tuyến trùng Meloidogyne sp. bằng cách tổng hợp enzyme thủy phân tác động lên da và trứng tuyến trùng (chitinase và protease) và các hợp chất thứ cấp ở dạng dễ bay hơi (Cyclohexasiloxane, dodecamethyl, Pentasiloxane, dodecamethyl-, 2,4-Dihydroxybenzoic acid, 3TMS derivative, 2,4,6-Cycloheptatrien-1-one, 3,5-bis-trimethylsilyl-.) nhưng chưa xác định được các hợp chất không có khả năng bay hơi. Vì vậy, nghiên cứu tiếp tục tiến hành phân tích, xác định các hợp chất thứ cấp tiềm năng không có khả năng bay hơi từ dịch nuôi cấy chủng RB.EK7 ở nội dung 3.2.3.
3.2.3. Phân tách, xác định cấu trúc và hoạt tính kháng tuyến trùng, kháng nấm của các hợp chất thứ cấp từ chủng vi sinh vật tuyển chọn
Các hợp chất kháng tuyến trùng (Bảng 3.31) là các hợp chất dễ bay hơi. Ngoài nhóm chất này, còn có nhiều các hợp chất thứ cấp tiềm năng cần tiếp tục nghiên cứu và làm sáng tỏ. Để tiếp tục nghiên cứu sâu hơn các hợp chất thứ cấp kháng tuyến trùng của vi khuẩn vùng rễ RB.EK7, dịch nuôi cấy được cô đặc, phân đoạn, tinh sạch và xác định cấu trúc hoá học bằng NMR, HMBC, HSQC.
3.2.3.1. Phân tách và xác định cấu trúc của các hợp chất
Dịch nuôi cấy của chủng RB.EK7 có bổ sung trứng và tuyến trùng tuổi 2 được cô đặc để tạo cao tổng. Cao này được lắc phân đoạn với các dung môi, thu hồi dung môi được các cao phân đoạn. Các cao chiết phân đoạn được tiến hành đánh giá khả năng ức chế tuyến trùng nhằm tìm ra phân đoạn có hoạt tính kháng tuyến trùng cao nhất. Kết quả ghi nhận ở bảng 3.32 cho thấy, hoạt tính kháng tuyến trùng cao nhất trên 90% ở cao chiết Ethyl acetate, cao chiết EA-Met (90:10) và cao chiết EA-Met (60:40) đạt từ 92,69% - 93,59%, kế đến là cao chiết chloroform và methanol đạt lần lượt là 86,88% và 86,49%.
Bảng 3.32 Khả năng ức chế tuyến trùng Meloidogyne sp. của cao chiết phân đoạn
Tỷ lệ tử vong (%) | |
1. Cao Chloroform | 86,88±2,67b |
2. Cao Ethyl acetate | 92,92±2,67a |
3. Cao EA -Met (90:10) | 93,59±1,28a |
4. Cao EA-Met (60:40) | 92,69±0,63a |
5. Cao Methanol | 86,49±3,16b |
6. Nước | 65,57±2,42c |
ĐC nước | 2,93±0,83e |
ĐC DMSO | 10,63±0,80d |
P | ** |
CV% | 3,06 |
*Ghi chú: ** Khác biệt có ý nghĩa thống kê ở mức p<0,01, các chữ cái giống nhau trên cùng một cột thể hiện sự khác biệt không có ý nghĩa thống kê theo trắc nghiệm phân hạng Duncan’s Multiple Rang Test.
Nhóm nghiên cứu của Gao và cộng sự (2016) đã tiến hành chiết phân đoạn dịch nuôi cấy của chủng Bacillus cereus với các dung môi hữu cơ khác nhau bao gồm ether dầu hỏa, chloroform, ethyl acetate, n-butyl alcohol và nước đồng thời đánh giá hoạt tính kháng tuyến trùng của các cao chiết [191]. Kết quả ghi nhận, cao chloroform có hoạt tính kháng tuyến trùng cao nhất với tỷ lệ tử vong tuyến trùng đạt 82,51%. Bên cạnh đó, hoạt tính kháng tuyến trùng cũng được ghi nhận ở phân đoạn tách với dung môi ethyl acetate và ete dầu hỏa [191].
Từ các cao chiết có hoạt tính kháng cao, nghiên cứu tiếp tục phân tách hợp chất kháng trên sắc ký cột hở và xác định cấu trúc của các hợp chất phân tách được. Kết quả phân tách được 5 hợp chất với tên lần lượt bao gồm VK01, VK02, VK03, VK04 và VK05 (Hình 3.20). Tuy nhiên, hai hợp chất VK02 và VK03 có cấu trúc không bền và dễ bị phân hủy. Do vậy, cấu trúc của 3 hợp chất VK01, VK04 và VK05 bằng phổ NMR, HMBC, HSQC lần lượt là dẫn xuất của Uracil, dẫn xuất của Thymine và hexahydropyrrolo [1,2-a] pyrazine-1,4-dione (Phụ lục 4).
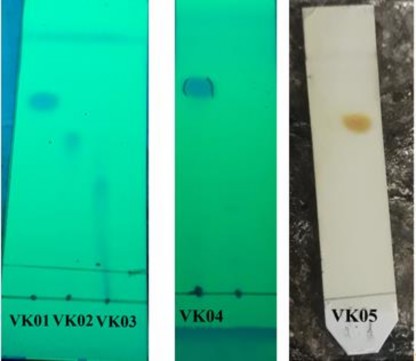
Hình 3.20. Sắc ký lớp mỏng các hợp chất phân tách
3.2.3.2. Đánh giá hoạt tính kháng tuyến trùng Meloidogyne sp. của các hợp chất đã phân tách
Sau khi phân tách được ba hợp chất bao gồm dẫn xuất của Uracil, dẫn xuất của Thymine và hexahydropyrrolo [1,2-a] pyrazine-1,4-dione, nghiên cứu tiến hành đánh giá hoạt tính kháng tuyến trùng thông qua tỷ lệ tử vong của tuyến trùng theo thời gian 3 giờ, 6 giờ, 9 giờ, 12 giờ và 24 giờ cũng như tỷ lệ nở của trứng tuyến trùng. Kết quả ghi nhận ở bảng 3.33, 3.34, 3.35.
Kết quả khảo sát nồng độ dẫn xuất của Uracil đến tỷ lệ tử vong tuyến trùng và tỷ lệ nở của trứng tuyến trùng ở bảng 3.33 cho thấy, có sự khác biệt có ý nghĩa thống kê giữa các nghiệm thức với p<0,01. Với nồng độ 2,5-20,0 mg/mL, dẫn xuất của Uracil làm tử vong 58,3-100% tuyến trùng tuổi 2 sau 24 giờ xử lý và tỷ lệ nở của trứng chỉ còn 13,7-59,7% so với 88,9% (ĐC nước). Oliveira và cộng sự (2014) đã tinh sạch được 3 hợp chất Uracil, dihidrouracil và 9H-purin trên cột sắc ký HPLC từ cao chiết diclomethane của dịch huyền phù vi khuẩn Bacillus cereus và Bacillus subtilis [192]. Uracil có thể gây tử vong tuyến trùng Meloidogyne exigua tuổi 2 đạt 38 % với nồng độ sử dụng là 500 µg/ml.
Bảng 3.33. Ảnh hưởng của dẫn xuất của Uracil đến tỷ lệ tử vong của tuyến trùng và tỷ lệ nở của trứng tuyến trùng
Tỷ lệ tử vong tuyến trùng (%) | Tỷ lệ nở của trứng tuyến trùng (%) | |||||
3 giờ | 6 giờ | 9 giờ | 12 giờ | 24 giờ | ||
2,5mg/ml | 18,86±1,35d | 28,88±0,93d | 39,26±0,46d | 49,42±0,52d | 58,36±0,34d | 59,71±1,10b |
5,0mg/ml | 25,42±1,01c | 38,90±0,12c | 49,38±0,33c | 58,69±0,37c | 69,53±0,13c | 35,06±1,75c |
10mg/ml | 37,35±1,07b | 49,48±0,93b | 64,59±0,82b | 74,56±0,32b | 89,36±0,41b | 23,58±1,32d |
20mg/ml | 47,77±1,17a | 65,16±1,03a | 78,65±0,68a | 88,24±0,78a | 100,00±0,00a | 13,79±1,01e |
ĐC DMSO | 0,00±0,00e | 0,00±0,00e | 0,00±0,00e | 1,37±0,26e | 3,26±0,17e | 87,63±0,41a |
ĐC nước | 0,00±0,00e | 0,00±0,00e | 0,00±0,00e | 0,00±0,00f | 0,00±0,00f | 88,93±0,49a |
P | ** | ** | ** | ** | ** | ** |
CV% | 4,48 | 2,37 | 1,13 | 0,94 | 0,38 | 2,32 |
*Ghi chú: ** Khác biệt có ý nghĩa thống kê ở mức p<0,01, các chữ cái giống nhau trên cùng một cột thể hiện sự khác biệt không có ý nghĩa thống kê theo trắc nghiệm phân hạng Duncan’s Multiple Rang Test.
Kết quả khảo sát nồng độ dẫn xuất của Thymine (một loại bazơ nitơ) đến tỷ lệ tử vong tuyến trùng và tỷ lệ nở của trứng tuyến trùng (Bảng 3.34) cho thấy, dẫn xuất của Thymine với nồng độ 2,5-20,0 mg/mL làm tử vong 67,99-100% tuyến trùng tuổi 2 sau 24 giờ xử lý và tỷ lệ nở của trứng chỉ còn 19,96-55,02% so với 92,36% (ĐC nước).
Bảng 3.34. Ảnh hưởng của dẫn xuất của Thymine đến tỷ lệ tử vong của tuyến trùng và tỷ lệ nở của trứng tuyến trùng
Tỷ lệ nở của trứng tuyến trùng (%) | ||||||
Nồng độ | 3 giờ | 6 giờ | 9 giờ | 12 giờ | 24 giờ | |
2,5mg/mL | 22,01±1,35d | 33,18±0,40b | 41,28±0,51b | 45,96±0,45d | 67,99±0,61d | 55,02±0,57c |
5,0mg/mL | 29,97±0,89c | 36,19±5,44b | 40,67±9,31b | 56,74±2,75c | 71,91±1,84c | 45,24±1,12d |
10mg/mL | 33,26±0,60b | 44,43±0,48a | 51,16±0,28a | 61,71±0,71b | 86,03±1,37b | 32,44±0,75e |
20mg/mL | 39,53±0,66a | 48,60±0,34a | 55,02±0,57a | 69,55±1,05a | 100,00±0,00a | 19,96±0,69f |
ĐC DMSO | 0,00±0,00e | 0,00±0,00c | 0,00±0,00c | 1,02±0,24e | 3,07±0,25e | 89,33±0,22b |
ĐC nước | 0,00±0,00e | 0,00±0,00c | 0,00±0,00c | 0,00±0,00e | 0,00±0,00f | 92,36±0,29a |
P | ** | ** | ** | ** | ** | ** |
CV% | 3,23 | 8,38 | 11,98 | 3,42 | 1,94 | 0,87 |
*Ghi chú: ** Khác biệt có ý nghĩa thống kê ở mức p<0,01, các chữ cái giống nhau trên cùng một cột thể hiện sự khác biệt không có ý nghĩa thống kê theo trắc nghiệm phân hạng Duncan’s Multiple Rang Test.