lượng và thành phần hoá học thịt. yijk = µ + Di + Sj + Di*Sj + εijk Trong đó
yijk : chỉ tiêu về sinh trưởng, năng suất thân thịt, chất lượng thịt và thành phần hoá học thịt
µ : trung bình quần thể
Di : ảnh hưởng của khẩu phần thứ ith (i = 4: ĐC, TN1, TN2 và TN3) Sj : ảnh hưởng của tính biệt thứ jth (j = 2: đực và cái)
Di*Sj: ảnh hưởng tương tác giữa khẩu phần và tính biệt εijk : sai số ngẫu nhiên
PHẦN 4. KẾT QUẢ VÀ THẢO LUẬN
4.1. GÂY ĐỘT BIẾN CHỦNG ASPERGILLUS NIGER A45.1 VÀ TỐI ƯU ĐIỀU KIỆN LÊN MEN CHỦNG ĐỘT BIẾN CHỌN LỌC CHO SINH TỔNG HỢP ĐA ENZYME (α-AMYLASE, GLUCOAMYLASE VÀ CELLULASE) CAO
4.1.1. Hoạt hóa giống
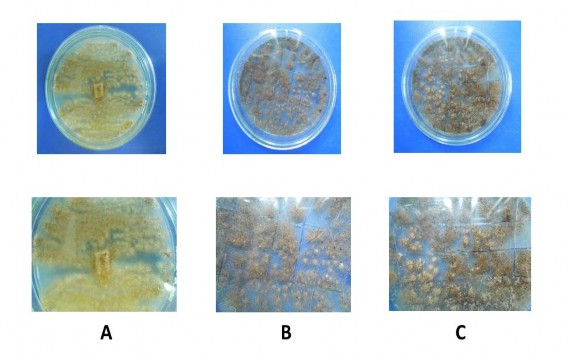
Từ chủng giống gốc Aspergillus niger A45.1 của Phòng Các chất chức năng sinh học, đã tiến hành hoạt hóa thành công giống trên môi trường PDA. Giống hoạt hóa có đặc điểm: khuẩn lạc tròn, màu sắc của khuẩn lạc thay đổi theo thời gian, tương ứng với các giai đoạn sinh trưởng và phát triển, từ trắng đến vàng và nâu, hệ sợi nấm mọc thưa, bào tử đen (Hình 4.1a).
Hình 4.1a. Giống hoạt hóa sau 5 ngày (A), 10 ngày (B) và 15 ngày (C)
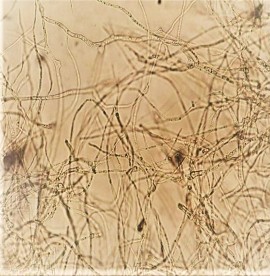
Quan sát hình thái của chủng Aspergillus niger A45.1 dưới kính hiển vi thấy, cành bào tử dạng bông, bào tử đính kép hai lớp, bào tử không nằm trong bọc bào tử, cuống sinh thể bình phình ra rõ rệt ở 2 đầu tạo bọng hình cầu (Hình 4.1b). Đặc điểm về định loại sinh học phân tử của chủng Aspergillus niger A45.1 được mô tả ở phụ lục 2.
| |
A. Cành bào tử nấm A13: 1: Cuống bào tử; 2: Cuống; 3: Bào tử đính. | B. Hệ sợi của chủng nấm A13 |
Có thể bạn quan tâm!
-

 Thành Tựu Về Cải Tiến Chủng Và Tối Ưu Môi Trường Lên Men Sản Xuất Amylase Và Cellulase
Thành Tựu Về Cải Tiến Chủng Và Tối Ưu Môi Trường Lên Men Sản Xuất Amylase Và Cellulase -
 Đặc Tính Và Vai Trò Của Các Vi Sinh Vật Sử Dụng Trong Lên Men Thức Ăn Chăn Nuôi
Đặc Tính Và Vai Trò Của Các Vi Sinh Vật Sử Dụng Trong Lên Men Thức Ăn Chăn Nuôi -
 Xử Lý Bã Sắn Thành Thức Ăn Chăn Nuôi Bằng Bằng Công Nghệ Đường Hóa Và Lên Men Đồng Thời
Xử Lý Bã Sắn Thành Thức Ăn Chăn Nuôi Bằng Bằng Công Nghệ Đường Hóa Và Lên Men Đồng Thời -
 Tính Ổn Định Của Các Chủng Đột Biến Chọn Lọc
Tính Ổn Định Của Các Chủng Đột Biến Chọn Lọc -
 Hoạt Tính Enzyme Của Chủng Aspergillus Niger Ga15 Khi Lên Men Xốp Ở Các Thời Gian Khác Nhau
Hoạt Tính Enzyme Của Chủng Aspergillus Niger Ga15 Khi Lên Men Xốp Ở Các Thời Gian Khác Nhau -
 Hàm Lượng Đường Khử, Tinh Bột Và Xơ Của Bã Sắn Ở Các Nồng Độ Enzyme Khác Nhau (N=3)
Hàm Lượng Đường Khử, Tinh Bột Và Xơ Của Bã Sắn Ở Các Nồng Độ Enzyme Khác Nhau (N=3)
Xem toàn bộ 197 trang tài liệu này.
Chú thích: Cuống bào tử và phần cuống dài từ phần sợi nấm (A); Sợi nấm có vách ngăn và bào tử nằm trong các sợi nấm (B)
Hình 4.1b. Hình ảnh hiển vi cấu tạo bào tử và sợi nấm của chủng nấm sợi
Aspergillus niger A45.1 (dưới vật kính 40X).
Chủng nấm sợi sau khi được hoạt hóa, tiến hành bảo quản trong tủ mát (4oC) để sử dụng trong các nghiên cứu tiếp theo, tiến hành cấy chuyển định kỳ, 7 ngày 1 lần.
4.1.2. Ảnh hưởng của tia UV và NTG đến khả năng sống sót của chủng nấm sợi Aspergillus niger A45.1
Để sản xuất enzyme thương mại, enzyme của nấm sợi được ưa chuộng hơn cả vì enzyme được tạo ra bởi các chủng nấm sợi hiệu quả hơn so với từ nấm men và vi khuẩn. Tuy nhiên, hầu hết các chủng dại mà có tiềm năng sử dụng trong quá trình lên men công nghiệp bắt buộc phải cải tiến để sự lên men có hiệu quả kinh tế. Đột biến là một công cụ thường được sử dụng để cải tiến chủng, được nhiều nhà khoa học sử dụng như một công cụ của kỹ thuật protein để đạt được những chủng có hiệu suất enzyme cao hoặc những đặc tính mong muốn. Gây đột biến bằng tia UV và hóa chất đã được sử dụng rộng rãi và phổ biến vì nó rất có hiệu quả và không đòi hỏi máy móc hay thiết bị phức tạp.
Để nâng cao khả năng sinh tổng hợp đa enzyme của chủng nấm sợi Aspergillus niger A45.1, tiến hành gây đột biến chủng bằng việc hòa tan bào tử vào dung dịch NTG và chiếu tia UV trong thời gian từ 0 đến 180 phút nhằm tạo ra những dòng biến thể mới có khả năng sinh tổng hợp enzyme cao hơn so với chủng gốc.
Kết quả cho thấy, tia UV và NTG có ảnh hưởng rõ rệt đến khả năng sống sót của các bào tử nấm do tia UV và NTG đã tạo nên các đột biến gây chết trên nấm sợi. Thời gian chiếu UV và xử lý NTG càng dài thì tỷ lệ sống sót của các bào tử càng giảm. Số lượng bào tử ở thời điểm chưa xử lý (0 phút) là 107 (bào tử/ml) sau đó giảm dần theo thời gian gây đột biến. Tỷ lệ (%) sống sót của các bào tử chỉ còn 26,73% sau 30 phút, 3,82% sau 60 phút và 0,4% sau 90 phút. Sau 120 phút xử lý, tỷ lệ sống sót bào tử chỉ với 0,12%. Từ sau 150 phút xử lý, lượng bào tử bị gây chết hoàn toàn (Hình 4.2 và Hình 4.3).
100
80
Bào tử sống sót (%)
60
40
20
0
0 30 60 90 120 150 180
Thời gian chiếu UV (phút)
Hình 4.2. Mối quan hệ giữa tỷ lệ (%) bào tử nấm Aspergillus niger A45.1 còn sống sót và thời gian chiếu tia UV và xử lý NTG
Tia UV đã được chứng minh là có khả năng gây chết và gây đột biến ở nhiều loài sinh vật, bao gồm cả nấm. Theo nghiên cứu của Reddy & cs. (2017), sau 60 phút xử lý bằng tia UV, khoảng 90% lượng bào tử của A.niger bị chết, chỉ còn khoảng 10% sống sót. Kết quả nghiên cứu của Shafique & cs. (2011) cũng cho biết, tỷ lệ chết của các bào tử nấm Trichoderma viride tăng khi tăng thời gian chiếu tia UV. Xu hướng tương tự cũng được phát hiện từ nghiên cứu này sau xử lý UV 90 và 120 phút, tỷ lệ sống sót còn lại tương ứng là 0,4% và 0,12%.
B
| C
| |
D
| E
| F.
|
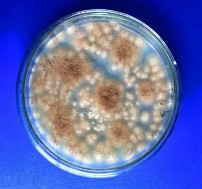
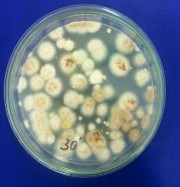
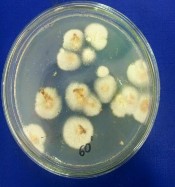
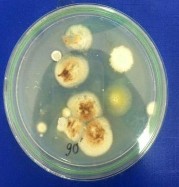
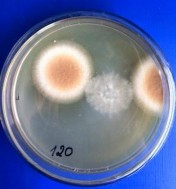
Hình 4.3. Các khuẩn lạc chủng Aspergillus niger A45.1 sau xử lý bởi tia UV và NTG (A: 0 phút; B: 30 phút; C: 60 phút; D: 90 phút; E: 120 phút; F: 180 phút)
Tia UV có thể tạo ra đột biến giữa 2 vòng pyrimidine để tạo nên 2 liên kết dimer giữa chúng. Qua quá trình sao chép ADN, cặp GC, GC (tự nhiên) sẽ tạo thành cặp AT, AT (đột biến) sau khi bị chiếu tia cực tím (Pathak & cs., 2015). NTG là một trong những hóa chất gây đột biến được sử dụng phổ biến trong phát triển chủng công nghiệp hiện nay, đây là tác nhân gây đột biến hiệu quả cao nhất đối với vi sinh vật thuộc lớp alkyl hóa. Dạng alkyl hóa gây đột biến bằng cách thêm nhóm methyl hoặc nhóm ethyl trên các bazơ, ở đó tạo sự thay đổi khi kết cặp của các bazơ trên sợi DNA. Cơ chế của sự đột biến này là đột biến điểm, làm biến đổi bazơ gây ra sự kết cặp sai. Khoảng 90% các đột biến do NTG gây ra là các đột biến điểm, thay thế cặp GC bằng cặp AT, một số rất ít các trường hợp dẫn đến dịch chuyển khung đọc hoặc mất đoạn (Vũ Văn Hạnh & cs., 2012). Tia UV và NTG có khả năng tạo các đột biến ngẫu nhiên trên vật liệu di truyền, tạo ra nguồn nguyên liệu biến đổi di truyền quan trọng để chọn lọc thu nhận dòng có nhiều đặc điểm tính trạng ưu việt hơn (Vũ Văn Hạnh & cs., 2012; Pathak & cs., 2015).
4.1.3. Hoạt tính enzyme của các các chủng đột biến
Các tế bào sống sót sau khi gây đột biến ở các khoảng thời gian khác nhau (liều gây đột biến khác nhau) được nhặt ngẫu nhiên, cấy chuyển sang các đĩa môi trường PDA, nuôi cấy ở 28oC trong 5 ngày. Tiến hành nghiên cứu khả năng thủy phân trực tiếp cơ chất tinh bột, xơ (CMC) trên đĩa thạch (PDA), chọn lọc được 20 chủng tiềm năng có khả năng sinh enzyme cao, lên men trên cơ chất xốp để xác định hoạt tính α-amylase, glucoamylase và cellulase của các chủng này.
Bảng 4.1. Hoạt tính enzyme của các chủng đột biến (n=3)
Chủng đột biến | Hoạt tính enzyme (U/g) (LSM)* | |||
Glucoamylase | α- amylase | Cellulase | ||
0 | A45.1 (Chủng dại) | 12,47fhi | 19,60cdef | 6,40bcdef |
GA11 | 10,88hi | 15,01defg | 4,26ef | |
GA12 | 14,41efh | 20,90cde | 6,31bcdef | |
30 | GA13 | 18,41cdef | 27,57bc | 4,66def |
GA14 | 21,41abcd | 32,57ab | 11,81a | |
GA15 | 27,41a | 38,68a | 12,16a | |
GA21 | 7,98hi | 10,18ghi | 6,08cdef | |
GA22 | 21,53abcd | 32,76ab | 8,81abcdf | |
60 | GA23 | 15,69def | 23,04cd | 12,68a |
GA24 | 21,31abcd | 32,40ab | 10,91ab | |
GA25 | 12,10fhi | 6,16hi | 8,38abcde | |
GA31 | 15,82edf | 8,02ghi | 7,99abcdef | |
GA32 | 20,38bcde | 10,30ghi | 9,42abcd | |
90 | GA33 | 22,43abcd | 11,33fghi | 6,55bcdef |
GA34 | 26,60ab | 13,41efgh | 5,77cdef | |
GA35 | 26,16ab | 13,19efgh | 9,77abc | |
GA41 | 22,82abc | 11,52fghi | 6,71cbdef | |
GA42 | 21,66abcd | 10,94ghi | 8,04bcdef | |
120 | GA43 | 6,21i | 3,21i | 6,99bcdef |
GA44 | 8,19hi | 4,17i | 3,41ef | |
GA45 | 16,42cdef | 8,28ghi | 4,51ef | |
SEM | 1,27 | 1,56 | 0,89 | |
P | <0,0001 | <0,0001 | <0,0001 |
*Trong cùng một cột, những giá trị LSM (trung bình bình phương nhỏ nhất) có các chữ cái khác nhau thì sai khác nhau có ý nghĩa thống kê (P<0,05).
Kết quả cho thấy, hoạt tính enzyme của các chủng đột biến có sự sai khác đáng kể (p<0,001). Hoạt tính enzyme glucoamylase cao thuộc về chủng GA15, GA14 (liều gây đột biến 30 phút), GA22, GA24 (liều gây đột biến 60 phút), GA33, GA34, GA35 (liều gây đột biến 90 phút), chủng GA42 (liều gây đột biến 120 phút) với hoạt tính glucoamylase trong khoảng 21,41 - 27,41 (U/g). Hoạt tính α-amylase cao ở các liều gây đột biến 30 phút với chủng GA15, GA14 và ở 60 phút với chủng GA22 và GA24, hoạt tính đạt được là 32,4 - 38,68 (U/g). Hoạt tính cellulase cao ở các liều gây đột biến 30, 60, 90 phút thuộc về các chủng GA15, GA14, GA23, GA24, GA25, GA31, GA32 và GA35, hoạt tính đạt được là 9,42 - 12,68 (U/g). Trong số các chủng đột biến kể trên, chủng GA15 là chủng có hoạt tính của cả 3 enzyme cao nhất. So với chủng dại, hoạt tính của α-amylase cao gấp 1,97 lần, glucoamylase cao gấp 2,2 lần, cellulase cao gấp 1,9 lần (Bảng 4.1).
Vi sinh vật có thể dễ dàng thao tác bằng cách sử dụng kỹ thuật di truyền hoặc các phương pháp khác để cải tiến chủng, tạo ra các đột biến và những thay đổi khác, nhờ đó việc sản xuất các enzyme có thể được tối ưu hóa. Trong hầu hết các trường hợp, tia UV gây ra những đột biến có hại nhưng đôi khi nó lại dẫn tới sự thích nghi của vi sinh vật với môi trường và làm cải thiện hiệu suất trao đổi sinh học (Irfan & cs., 2011). Việc chiếu tia UV được tìm thấy là cách tốt nhất để cải thiện các chủng như Aspergillus niger để sản xuất tối đa nhiều loại enzyme (Olukunle & Ajayi, 2018). Olukunle & Ajayi (2018) tiến hành cải tiến chủng A. flavus và A.niger bằng gây đột biến với tia UV ở bước sóng 240nm trong 10, 20 và 30 phút, kết quả cho thấy hoạt độ của amylase được tạo ra bởi các chủng nấm dao động từ 2,85 U/ml/phút đến 3,263 U/ml/phút. Trong đó, các chủng đột biến của A. niger ở liều tiếp xúc tia UV trong 10 phút có khả năng sản xuất amylase cao nhất. Raju & cs. (2012) đã nghiên cứu sự cải tiến của A. niger cho sản xuất glucoamylase bằng tác nhân vật lý (UV) và hóa học (Ethyl methyl sulphonate và ethidium bromide) và báo cáo rằng các chủng đột biến của A. niger có khả năng sản xuất glucoamylase cao hơn so với chủng dại. Vu & cs. (2010) đã nghiên cứu việc sản xuất cao sản enzyme phân hủy tinh bột sống (RSDE) bằng việc gây đột biến lặp lại chủng nấm sợi Aspergillus niger và nhận thấy rằng sự sản sinh RSDE được cải thiện cao hơn 2 lần so với chủng dại. Irfan & cs. (2011) đã tiến hành cải thiện sự sản xuất enzyme của chủng A.niger bằng gây đột biến với tia UV. Kết quả cho thấy, tia UV đã gây ra các đột biến và tăng cường hoạt tính của CMCase gấp 2 lần, của Fpase lên đến 3 lần so với chủng dại. Đối với enzyme avicelase, xylanase và sản xuất sinh khối nấm thì bức xạ tia UV chỉ ảnh hưởng nhẹ so với dòng dại. Theo Agrawal &
cs. (1999) tia UV là một chất gây đột biến mạnh. Tia UV được tìm thấy là tốt nhất trong việc cải thiện các chủng như A.niger cho sự sản xuất tối đa các loại enzyme khác nhau. Chủng A. niger đột biến bởi tia UV có khả năng sản xuất lipase bằng 156% so với chủng dại. Chủng A.niger UAM-được xử lý bằng UV tăng sản xuất memicellolytic và cellulolytic (Nicolás-Santiago & cs., 2006) cho biết các chủng đột biến từ A. niger UAM - GS1 được tạo ra bằng chiếu tia UV đã tăng đáng kể mức sản xuất các enzyme mannanase, xylanase và cellulase của chúng so với chủng dại ban đầu, cải thiện đáng kể các ứng dụng công nghiệp tiềm năng của chúng.
Một số kết quả nghiên cứu đã chỉ ra rằng, các chủng đột biến do chiếu tia UV không có sự thay đổi nào trong hệ gen của chúng, khả năng tăng hoạt tính enzyme của những chủng này là do những thay đổi có thể xảy ra trong vùng promoter của các gen mã hóa cho các enzyme này. Tia phóng xạ có thể phá hủy sự điều hòa phiên mã của mARN tương ứng của enzyme, dẫn tới sản xuất enzyme (Nicolás-Santiago & cs., 2006; Li & cs., 2010; Singh & cs., 2013).
NTG cũng được coi là một chất gây đột biến hóa học hiệu quả, đây là chất alkyl hóa mạnh, làm đứt mạch ADN (Huang & cs., 2019). Vu & cs. (2011) cải tiến chủng nấm sợi Aspergillus niger SU14 bằng tia Co60, UV và NTG đã lựa chọn được một chủng đột biến có khả năng sản sinh cellulase tăng gấp 2,2 lần so với chủng dại. Vu & cs. (2010) cho biết, sự kết hợp các nhân tố đột biến như tia γ (Co60), UV và NTG đã cho hiệu quả cao hơn trong việc gây đột biến các chủng nấm sợi để làm tăng khả năng sản xuất RSDE và lên men xốp ở điều kiện tối ưu đã làm tăng hiệu quả sản xuất RSDE cao gấp 19,23 lần so với kiểu dại ở cùng điều kiện.
Nhiều nghiên cứu chỉ ra rằng, việc kết hợp các tác nhân gây đột biến hóa học và vật lý trong cải tiến chủng đã làm tăng hoạt tính của một số enzyme của nhiều chủng vi sinh vật. Zhao & cs. (2014) nghiên cứu sự kết hợp của tia UV và tia Gamma Co60 trong gây đột biến A.niger để tăng cường hoạt tính của glucoamylase. Abdullah & cs. (2013) tiến hành gây đột biến chủng Aspergillus oryrae để tăng cường sản sinh α-amylase bằng tia UV và EMS, kết quả cho thấy hoạt tính của α-amylase của chủng đột biến tăng 2,1 lần so với chủng dại. Karanam & cs. (2008) cho biết, khi kết hợp tia UV, HNO2 và NTG đã làm tăng hoạt tính lipase của Aspergillus japonicus MTCC 1975 lên 276% so với chủng bố mẹ. Nadir & cs. (2013) cũng báo cáo rằng sự kết hợp của các nhân tố gây đột biến để cải tiến chủng Leuconostoc mesenteries đã làm tăng hoạt tính α-amylase gấp 7 lần và của glucoamylase gấp 3 lần so với chủng bố mẹ.