tế bào apoptotic đã được quan sát thấy ở chuột không có MBL (tức là loại bỏ kép MBLA/C). Tuy nhiên, bất chấp sự lưu thông kéo dài của các tế bào apoptotic, những con chuột này khi già đi đã không phát triển bất kỳ dấu hiệu nào của bệnh tự miễn dịch . Những dữ liệu này cho thấy MBL có vai trò trong việc loại bỏ các tế bào đang chết, nhưng cũng gợi ý rằng để phát triển bệnh tự miễn dịch đòi hỏi nhiều yếu tố nữa bên cạnh sự ứ đọng tế bào chết theo chương trình.
Các nghiên cứu đã chỉ ra rằng MBL có thể liên kết với các tế bào bị
stress và bị
tổn thương do thiếu máu cục bộ
sau chấn thương. Sự
lắng
đọng MBL bắt đầu kích hoạt bổ thể theo cách độc lập với C1q và điều này góp phần gây ra tổn thương tế bào chủ . Một số kháng nguyên mới có khả năng được phát hiện trong tổn thương do thiếu máu cục bộ. Cytokeratin1 đại diện cho các phân tử là phối tử của MBL trong điều kiện tế bào stress và tổn thương . Ngoài ra, MBL có khả năng liên kết với một số dòng tế bào khối u của người đã được biến đổi có các kiểu glycosyl hóa theo cơ chế như Lewis A và Lewis B (Lea – Leb). Những cấu trúc này được biểu hiện trên các phân tử glycoprotein của các dòng tế bào có nguồn gốc từ người tạo thành ung thư biểu mô tuyến ruột kết và ung thư biểu mô đại trực tràng
, . Ở những dòng tế bào này, chuỗi nặng CD26 và CD98 đã được xác định là các glycoprotein có chứa cấu trúc LeaLeb là các phối tử cho MBL . Hơn nữa, MBL có liên quan đến việc liên kết với các đồng phân của các kháng thể được tìm thấy trong viêm khớp dạng thấp .
Vai trò của MBL trong hoạt hóa hệ thống bổ thể
Có thể bạn quan tâm!
-
 Giá Trị Phân Biệt 2 Nhóm Bệnh Dhf Và Dws Của Protein 83
Giá Trị Phân Biệt 2 Nhóm Bệnh Dhf Và Dws Của Protein 83 -
 Cơ Chế Bệnh Sinh Của Sốt Xuất Huyết Dengue
Cơ Chế Bệnh Sinh Của Sốt Xuất Huyết Dengue -
 Nghiên cứu tính đa hình gen MBL2, FCN2 và nồng độ protein MBL, Ficolin-2 ở bệnh nhân sốt xuất huyết Dengue - 5
Nghiên cứu tính đa hình gen MBL2, FCN2 và nồng độ protein MBL, Ficolin-2 ở bệnh nhân sốt xuất huyết Dengue - 5 -
 Vai Trò Của Mbl, Ficolin2 Và Tính Đa Hình Gen Mbl2, Fcn2 Trong Sốt Xuất Huyết Dengue
Vai Trò Của Mbl, Ficolin2 Và Tính Đa Hình Gen Mbl2, Fcn2 Trong Sốt Xuất Huyết Dengue -
 Chỉ Tiêu Về Lâm Sàng, Cận Lâm Sàng
Chỉ Tiêu Về Lâm Sàng, Cận Lâm Sàng -
 Xác Định Nồng Độ Ficolins2 Và Mbl Trong Huyết Thanh
Xác Định Nồng Độ Ficolins2 Và Mbl Trong Huyết Thanh
Xem toàn bộ 189 trang tài liệu này.
MBL khởi động con đường hoạt hóa bổ thể lectin bằng cách liên kết với các serin protease tạo ra các MASP , . Có bốn MASP đã được tìm thấy
là MASP1, 2, 3 và sMAP ∕MAP19. MASP1 và MASP2 được mã hóa bởi các gen riêng biệt trong khi MASP3 và sMAP là sản phẩm của sự nối ghép
thay đổi của các gen MASP1 và MASP2 tương
ứng. Chỉ
MASP1 và
MASP2 có miền serin protease . MBL và MASPs hiện diện dưới dạng phức hợp trong tuần hoàn . Khi MBL liên kết với các phối tử trên bề mặt vi sinh vật sẽ tự động kích hoạt MASP2, cho phép phân cắt C4 và C2 để tạo thành C3 convertase cổ điển C4bC2a. Liên kết MBL MASP xảy ra thông qua vùng collagen của MBL . Cần lưu ý rằng các phân tử trong huyết thanh chứa cấu trúc giống collagen khác như Ficolin khi liên kết với một số loại đường nhất định cũng có khả năng kích hoạt bổ thể theo con đường lectin thông qua MASP , . Những kết quả này chỉ ra rằng cấu trúc giống collagen
đóng một vai trò quan trọng trong việc kích hoạt bổ lectin thông qua liên kết với MASP.
1.4.2. Gen MBL2 và tính đa hình gen MBL2
thể
theo con đường
Việc phát hiện ra khiếm khuyết thực bào là do thiếu hụt MBL đã khởi
đầu một nỗ lực giải quyết cơ chế phân tử đằng sau sự thiếu hụt MBL .
Giải trình tự các exon của gen MBL2 từ ba đứa trẻ có nồng độ MBL thấp trong huyết thanh và thiếu hụt khả năng thực bào và bị tái phát nhiễm trùng tiết lộ rằng mỗi đứa trẻ mang một đột biến điểm tại codon 54 trong exon 1 gây ra sự thay thế glycine bằng acid aspartic (GGC thành GAC) (alen B, alen bình thường được đặt tên là A) (Hình 1.8) . Nghiên cứu các thành viên trong gia đình của các nhóm đã gợi ý sự thừa kế ưu thế của NST, thường là nồng độ MBL thấp vì nồng độ protein trung bình giảm khoảng 10 lần ở cá thể có kiểu gen dị hợp tử. Tác động giảm nồng độ MBL huyết thanh của alen
B là do sự lắp ráp sai cấu trúc chuỗi xoắn ba MBL.Nếu chuỗi biến thể
tham gia vào quá trình lắp ráp chỉ 1/8 của cấu trúc bộ ba cơ bản, về mặt
thống kê sẽ tránh sự hiện diện của một polypeptid biến thể, trong khi bảy phần còn lại sẽ chứa một hoặc nhiều polypeptid bị lỗi và do đó làm phát
sinh một phân tử
không
ổn định. Tuy nhiên, alen
B lại xuất hiện thường
xuyên một cách đáng ngạc nhiên ở người da trắng khỏe mạnh (0,13) và
người nhập cư từ Đông Greenland, trong khi hiếm gặp ở Đông Phi .
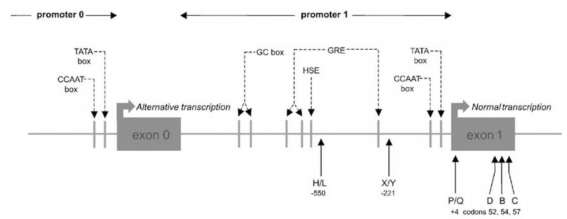
Hình 1.8: Đa hình gen MBL2
*Nguồn: theo Garred P. và cs (2006)
Một đột biến khác ở codon 57 (GGA thành GAA) khiến glycine được thay thế bằng acid glutamic (alen C) được tìm thấy ở các cá thể từ Tây Phi (Hình 1.8) . Alen C rất phổ biến và được tìm thấy với tần số cao đáng kể
(0,29). Tuy nhiên, alen C được tìm thấy với tần số
rất thấp
ở người da
trắng và nó không có ở người châu Á và người da đen thuần chủng. Giống với alen B, alen C cũng ảnh hưởng đến nồng độ MBL huyết thanh. Một đột
biến ở
codon 52 (CGT thành TGT) khiến arginine được thay thế
bằng
cysteine (alen D) (Hình 1.8) . Alen này có tần số 0,05 ở cả người Đông Phi và người da trắng. Tuy nhiên, giống alen C, nó hầu như không có ở người châu Á và ở người Inuits và thậm chí ở người Tây Phi. Alen D cũng có ảnh
hưởng làm giảm nồng độ
MBL trung bình ở dị
hợp tử A/D, nhưng
ảnh
hưởng ít hơn và dải nồng độ MBL trong huyết thanh rộng hơn so với hai
alen còn lại. Tuy nhiên, ở thể đồng hợp tử D/D hoặc ở thể dị hợp tử kết
hợp (B/D và C/D) nồng độ MBL huyết thanh giảm tương tự như đối với
các kiểu gen B/B, C/C và B/C. Vì tác động lên MBL huyết thanh khá giống nhau, để thuận tiện trong phân tích thống kê, các biến thể cấu trúc B, C và D thường được gộp chung là alen O .
Vẫn còn một sự
khác biệt khá lớn về
nồng độ
MBL trong huyết
thanh ở những người không mang alen O. Điều này có thể được giải thích
bởi các đa hình được tìm thấy trong vùng promoter của gen MBL2 (Hình
1.8) , . Đặc biệt là hai điểm đa hình nằm ở các vị trí 550 (biến thể H/L) và
221 (biến thể X/Y), cả hai đều là sự thay thế nucleotide G thành C được chứng minh là có vai trò quan trọng . Dựa trên vị trí các đa hình trên nhiễm
sắc thể có thể xác định các kiểu gen đơn bội phổ biến sau: HYA, LYA và
LXA trên nền nhiễm sắc thể bình thường và HYD, LYB và LYC trên nhiễm sắc thể mang alen biến thể cấu trúc. Các kiểu gen đơn bội HYA, LYA, LXA tương quan khá tốt với nồng độ MBL huyết thanh cao, trung bình và thấp tương ứng. Tuy nhiên, kiểu gen đơn bội LYA có thể được chia nhỏ hơn nữa thành các kiểu gen đơn bội khác khi đa hình (P/Q) nằm trong phần 5’UTR
của exon 1 (vị trí +4) được đề cập đến , . Như vậy, kiểu LY bao gồm các
kiểu gen đơn bội sau: LYPA, LYPB, LYQA và LYQC. Cũng có thể quan sát thấy sự khác biệt về nồng độ trong huyết thanh giữa các kiểu gen đơn bội LYPA và LYQA cho thấy rằng mỗi kiểu gen đơn bội của promoter được xác định có liên quan đến các nồng độ MBL khác nhau trong huyết thanh .
Trong một phân tích chức năng vùng promoter, người ta thấy rằng ba
kiểu gen đơn bội
HY, LY và LX tương quan với mức độ
hoạt động cao,
trung bình và thấp của vùng promoter, phù hợp với nồng độ MBL huyết
thanh . Các kiểu gen đơn bội của gen MBL2 được bảo tồn rất tốt trong tất
cả các quần thể
được nghiên cứu và sự
khác biệt về
tần số
có thể
giải
thích sự khác biệt nồng độ MBL huyết thanh giữa các chủng tộc khác nhau. Mặc dù sự thay đổi nồng độ MBL huyết thanh có thể được giải thích bởi
các kiểu gen đơn bội, vẫn không thể giải thích được một số trường hợp
thay đổi nồng độ MBL huyết thanh. Điều này cho thấy rằng còn có các yếu tố di truyền khác nữa ảnh hưởng đến nồng độ MBL trong huyết thanh .
Việc giải trình tự của vùng 10 kb bao gồm toàn bộ gen MBL2 (gồm cả intron) đã được thực hiện ở các cá nhân đại diện cho bốn nhóm dân tộc chính của Mỹ cũng như theo dõi ở người da trắng Hà Lan . Kết quả có 87 vị trí đa hình đã được phát hiện. Những nghiên cứu này đã xác nhận sự bảo
tồn mạnh mẽ
của bảy dạng đơn bội
MBL2 cổ
điển. Tuy nhiên, các
ước
tính về sự mất cân bằng liên kết trên gen MBL2 chỉ ra rằng gen được chia thành hai vùng với điểm nóng tái tổ hợp có thể xảy ra ở đầu 3’. Do đó, rõ ràng có sự hiện diện của các đa hình khác nữa trong vùng lân cận của gen MBL2 hoặc ở vùng intron, có thể xuất hiện cùng các dạng đơn bội cổ điển khác nhau.
Việc giải trình tự gen MBL2 từ các cá thể mang các kiểu gen khác
nhau đã cho thấy sự tồn tại của năm loại thay thế hoặc xóa bỏ bazơ trong vùng promoter của các kiểu gen đơn bội LYQA và LYQC khác với các kiểu gen đơn bội LYPA và LYPB . Những đa hình này phân biệt kiểu gen đơn bội LYP với kiểu gen đơn bội LYQ. Các kiểu gen đơn bội trung gian chưa được mô tả ở người nhưng trình tự sắp xếp của các loài linh trưởng đã cho thấy các dạng trung gian . Những kết quả này chỉ ra rằng các kiểu gen đơn bội
trung gian có thể
đã bị
mất trong quá trình tiến hóa và kiểu gen đơn bội
LYQA và LYPA của con người có thể đã phát sinh mới (Hình 1.9). Cấu trúc exon của gen MBL2 được bảo tồn cực kỳ tốt giữa con người và động vật
linh trưởng bậc cao . Đặc biệt, vùng giống collagen ở người, tinh tinh và khỉ đột ở cấp độ acid amin giống hệt nhau và sự khác biệt trong toàn bộ protein nhỏ hơn 1%. Những khác biệt nhỏ về acid amin này không dẫn đến thay đổi cấu trúc hoặc chức năng của MBL .
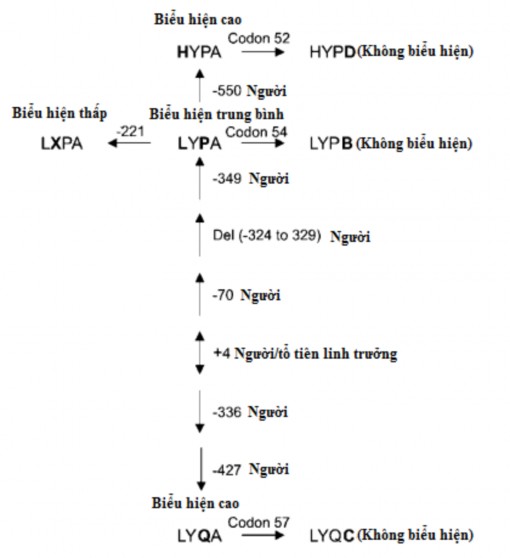
Hình 1.9: Cây tiến hóa MBL2
*Nguồn: theo Garred P. và cs (2006)
1.5. Protein Ficolin2 và gen FCN2
1.5.1. Protein Ficolin2
1.5.1.1. Cấu trúc protein Ficolin2
Ficolin2 hay LFicolin ở người là một tập hợp (có thể là 12 tiểu đơn
vị) của các tiểu đơn vị có kích thước 35 kDaa. Mỗi tiểu đơn vị bao gồm
vùng tận cùng N ngắn gồm 13 acid amin giàu cysteine, vùng giống collagen
ở giữa với 2 bộ ba GlyXY, tiếp theo là vùng trống khoảng sáu gốc acid
amin và phần lặp lại 15 lần GlyXY, một vùng cổ ngắn gồm 9 acid amin và vùng giống fibrinogen hình cầu gồm 209 acid amin . Cấu trúc oligomeric
của Ficolin2 được hình thành là do sự liên kết chéo của ba tiểu đơn vị
thông qua cầu nối disulfua trong vùng đầu cuối N (Hình 1.10). Vùng giống
fibrinogen tạo thành cấu trúc hình cầu giống như vùng nhận biết
carbohydrate của MBL. Vùng giống collagen tập hợp các tiểu đơn vị thành bộ ba, và vùng đầu cuối N lắp bốn bộ ba thành một cấu trúc gồm 12–24 tiểu đơn vị. Hai gốc cysteine trong vùng tận cùng N được cho là có vai trò
trung gian trong phản ứng trùng hợp bằng liên kết disulfide. Ohashi .T và
Erickson H.P. đã đề xuất một mô hình trong đó các liên kết disulfide Cys24
– Cys24 đối xứng giữa các bộ ba là nền tảng cho sự trùng hợp . Cấu trúc tổng thể của Ficolin2 trông giống như một "bó hoa" (Hình 1.10).
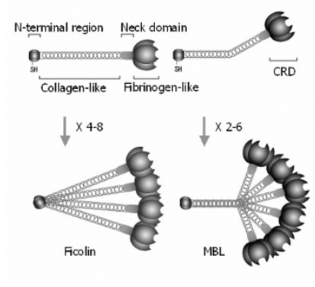
Hình 1.10: Cấu trúc đa bậc của protein Ficolin và MBL
*Nguồn: theo Endo Y. và cs (2006)
1.5.1.2. Chức năng của protein Ficolin2 Gắn với carbohydrate
Ficolin2 liên kết với GlcNAc trong chuỗi GlcNAcBSA và chuỗi oligosaccharide loại phức hợp có gốc GlcNAc liên kết với lõi trimannosyl, nhưng không liên kết với mannoseBSA hoặc các mannose, oligosaccharides dạng lai . Không giống như MBL, Ficolin2 có thể liên kết với GlcNAc bên
cạnh galactose không hoạt động trong oligosaccharide. Khả năng liên kết
với carbohydrate của Ficolin2 được thực hiện bởi vùng giống fibrinogen. Vùng giống fibrinogen của Ficolin2 cho thấy sự tương đồng với nửa đầu C của chuỗi fibrinogen β và γ. Trước đây, Doolittle đã chỉ ra rằng cấu trúc
chính của fibrinogen có liên quan đến CRD của MBL, đặc biệt là về sự
phối hợp của các gốc acid amin tích điện cần thiết cho sự liên kết với các phân tử đường .
Gắn với vi khuẩn