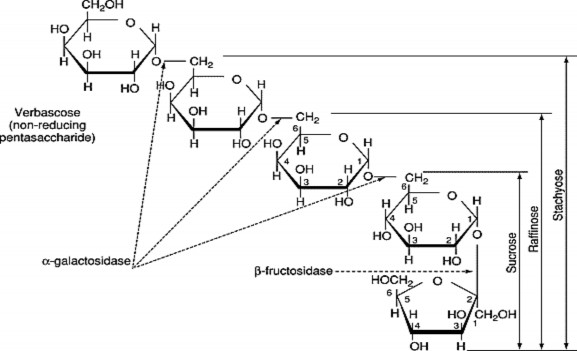
Hình 1.3. Cấu tạo và mối liên hệ của các đường thuộc họ Raffinose
O -α- d -Galactopyranosyl-(1→ 6) - [ O -α- d -galactopyranosyl- (1→ 6)] 2 - O -α- d -glucopyranosyl- (1→ 2) β- d -frutofuranoside. Các mũi tên chỉ ra
các điểm bị thủy phân bởi các enzyme [12].
RFOs là chất lưu trữ phổ biến có trong hạt đậu tương, nó ảnh hưởng đến khả năng giữ ẩm của hạt và khả năng chịu stress của cây [13]. Một số nghiên cứu gần đây cho thấy trong hạt đậu tương của các giống đang được gieo trồng có chứa hàm lượng cao stachyose và raffinose [14]. Bên cạnh đó, một vài nghiên cứu về hạt đậu tương có lượng hàm stachyose và raffinose thấp hoặc raffinose tổng thể (RFOs) lại thấp hơn so với cây hoang dại, mà không dẫn tới sự khác biệt đáng kể về khả năng chống chịu stress hoặc khả năng nảy mầm của hạt [15]. Trên các giống đậu tương thuần chủng thành phần chính trong RFOs là stachyose (30-60 g/1 kg), tiếp đến là raffinose (2-9 g/1 kg) [16]. Nghiên cứu của Kumar cũng chỉ ra rằng trong hạt đậu tương có chứa từ 6,4-25,3 mM/kg raffinose và 20,9-71,0 mmol/kg stachyose [17]. Ở hạt trưởng thành RFOs chủ yếu dạng stachyose, các hợp chất này có nguồn gốc từ sucrose có tác dụng tích cực đến quá trình chuyển hóa năng lượng. Tuy nhiên đây là nhóm đường khó tiêu hóa đối với người và động vật dạ dày đơn [10].
Các nghiên cứu vừa qua cho thấy RFOs là nguồn năng lượng ảnh hưởng đến quá trình nảy mầm của hạt, việc ức chế chuyển hóa RFOs làm giảm đáng kể khả năng nảy mầm của hạt [18]. Hàm lượng stachyose trong hạt tỉ lệ thuận với khả năng chịu
ẩm của hạt đậu tương, đối với hạt non bị khô nước sẽ chậm làm tăng tích lũy RFOs [19]. Những oligosaccharide khó tiêu hóa này thông qua hệ tiêu hóa đi từ qua dạ dày đến ruột non, sau đó chúng được lên men bởi các vi sinh vật kỵ khí cư trú trong ruột già. Quá trình lên men tạo ra khí CO2 và CH4 làm rối loạn hoạt động tiêu hóa dẫn đến chứng đầy hơi và tiêu chảy ở động vật dạ dày đơn [20].
Mối quan tâm đến các yếu tố gây ra sự thay đổi này bắt nguồn từ ý nghĩa nông nghiệp của RFOs. Do đặc tính khó chuyển hóa trong hệ tiêu hóa của người và động vật dạ dày đơn nên việc loại bỏ RFOs trong hạt của cây trồng vẫn đang được nghiên cứu tích cực [21].
Có thể bạn quan tâm!
-

 Nghiên cứu ứng dụng công nghệ CRISPR/Cas9 trong tạo đột biến gen GmGOLS03, GmGOLS19 trên cây đậu tương Glycine max L. Merrill nhằm giảm lượng đường họ Raffinose trong hạt - 1
Nghiên cứu ứng dụng công nghệ CRISPR/Cas9 trong tạo đột biến gen GmGOLS03, GmGOLS19 trên cây đậu tương Glycine max L. Merrill nhằm giảm lượng đường họ Raffinose trong hạt - 1 -
 Nghiên cứu ứng dụng công nghệ CRISPR/Cas9 trong tạo đột biến gen GmGOLS03, GmGOLS19 trên cây đậu tương Glycine max L. Merrill nhằm giảm lượng đường họ Raffinose trong hạt - 2
Nghiên cứu ứng dụng công nghệ CRISPR/Cas9 trong tạo đột biến gen GmGOLS03, GmGOLS19 trên cây đậu tương Glycine max L. Merrill nhằm giảm lượng đường họ Raffinose trong hạt - 2 -
 Tình Hình Sản Xuất Và Chọn Tạo Giống Đậu Tương
Tình Hình Sản Xuất Và Chọn Tạo Giống Đậu Tương -
![Quá Trình Hoạt Động Của Cơ Chế Crispr/cas Ở Vi Khuẩn Chống Lại Sự Xâm Nhập Của Dna Ngoại Lai [51].](https://tailieuthamkhao.com/uploads/2023/02/20/nghien-cuu-ung-dung-cong-nghe-crispr-cas9-trong-tao-dot-bien-gen-gmgols03-5-1-120x90.jpg) Quá Trình Hoạt Động Của Cơ Chế Crispr/cas Ở Vi Khuẩn Chống Lại Sự Xâm Nhập Của Dna Ngoại Lai [51].
Quá Trình Hoạt Động Của Cơ Chế Crispr/cas Ở Vi Khuẩn Chống Lại Sự Xâm Nhập Của Dna Ngoại Lai [51]. -
 Nghiên Cứu Biểu Hiện Gen Và Chỉnh Sửa Hệ Gen Trên Rễ Tơ Đậu Tương
Nghiên Cứu Biểu Hiện Gen Và Chỉnh Sửa Hệ Gen Trên Rễ Tơ Đậu Tương -
 Chuyển Gen Tạo Cây Đậu Tương Mang Cấu Trúc Crispr/cas9
Chuyển Gen Tạo Cây Đậu Tương Mang Cấu Trúc Crispr/cas9
Xem toàn bộ 122 trang tài liệu này.
Raffinose
Raffinose (RFO) là một trisaccharide gồm galactose, glucose và fructose. Chúng có thể được tìm thấy trong đậu, bắp cải, cải bruxen, bông cải xanh, măng tây, các loại rau khác và ngũ cốc nguyên hạt. RFO có khối lượng phân tử cao hơn Stachyose, Verbascose và Ajugose Raffinose, có thể được thủy phân thành D- galactose và sucrose bởi enzyme α-galactosidase (α-GAL), một loại enzyme không có trong đường tiêu hóa của con người [22]. Các nhà khoa học cho rằng RFO được xem như tác nhân bảo vệ phôi hạt trong quá trình tạo độ ẩm và bảo quản hạt ở trạng thái khô [13], tuy nhiên không có bằng chứng về mối quan hệ giữa sự tích tụ của chúng và khả năng chịu ẩm. RFO thường tích tụ trong các mô sinh dưỡng như một kháng thể của thực vật giúp cây chống lại stress sinh học và phi sinh học [23].
Stachyose
Một trong những tetrasaccharide dồi dào trong thực vật là stachyose. Stachyose bao gồm bốn đơn vị đường: hai đơn vị α-D-galactoza, một đơn vị α-D- glucoza và một đơn vị β-D-fructose lần lượt liên kết như sau gal(α1→6) gal (α1→6) glc (α1↔2β) fru. Stachyose lần đầu tiên được phân lập từ thân rễ của Stachys tuberifera, cùng tồn tại với RFO và các oligosaccharide liên quan khác trong các cơ quan khác nhau của nhiều loài thực vật. Trong cây thân gỗ, họ bầu bí và cây họ đậu loại đường này tồn tại trong không bào, rễ, hạt... mang chức năng dự trữ và vận chuyển trong cây.
Các tế bào trung gian là nơi chủ yếu tổng hợp RFO và stachyose. Trong
lá Catalpa và Buddleia, stachyose được tổng hợp từ sự kết hợp của RFO và galactinol thông qua quá trình quang hợp. Enzyme raffinose synthase (RS) không thể tổng hợp stachyose, một loại enzyme cụ thể tồn tại trong nguồn thực vật đã tổng hợp tetrasaccharide này. Enzyme này được phân lập từ một số nguồn và được tìm thấy để xúc tác phản ứng sau [24]:
[14C] Galactinol + raffinose →[14C] stachyose + myo-inositol
Tuy nhiên, phản ứng chuyển vị có thể thuận nghịch tự do, nhưng quá trình tổng hợp stachyose được ưu tiên [25]:
Stachyose + [14C] raffinose ⇌ [14C] stachyose + raffinose
Ở thực vật, stachyose được tổng hợp trong quá trình phát triển của lá và quá trình tổng hợp đạt mức cao nhất ở giai đoạn lá trưởng thành. Stachyose sẽ vận chuyển đến tất cả các bộ phận khác trong cây thông qua mạch rây (libe), ngoại trừ lá trưởng thành khác. Sự tổng hợp stachyose xảy ra ngay cả ở những lá già và một phần bị vàng.
Trong hạt, stachyose được chuyển hóa trong quá trình nảy mầm. Ở những cây cứng mùa đông, stachyose và các oligosaccharide liên quan tạo nên sự cứng cáp cho cây; hàm lượng có sự thay đổi theo mùa về kích thước vách tế chúng. Stachyose chủ yếu được sử dụng làm chất tạo ngọt hoặc sử dụng với tính chất, chức năng của một oligosaccharide. Con người không tiêu hóa được stachyose hoàn toàn.
Verbascose
Verbascose là một pentasaccharide có mặt trong tầng libe và trong các cơ quan lưu trữ của nhiều loài thực vật, đặc biệt là trong hạt của cây họ đậu [26]. Nhóm đường này có rất ít trong hạt nên nghiên cứu về chức năng sinh lý của verbascose trong hạt cũng hạn chế. Verbascose có thể hoạt động như một chất bảo vệ trong quá trình trưởng thành hạt bị khô hạn hoặc là nguồn dự trữ carbon cho quá trình hạt nảy mầm [27]. Hàm lượng của verbascose trong hạt có thể biến đổi mạnh, cụ thể nghiên cứu về sự thay đổi hàm lượng verbascose ở đậu faba (Vicia faba) thay đổi từ 1,1% đến 9,9% chất khô của hạt [28]. Trong một cuộc khảo sát về kiểu gen của hạt đậu (Pisum sativum), các giá trị về hàm lượng verbascose dao động từ mức không thể phát hiện đến 3,1% chất khô.
Nghiên cứu ban đầu chỉ ra rằng sinh tổng hợp verbascose tiến hành bằng cách chuyển một đơn vị galactose từ galactinol [O ‐ α ‐ d ‐ galactopyranosyl‐ (1 → 1) ) l ‐ myo ‐ inositol] thành stachyose. Việc phân lập được enzyme verbascose synthase
(VBS) từ hạt đậu của các nhà khoa học đã chứng minh đây là quá trình tổng hợp stachyose đa chức năng [29]. Enzyme hoạt động theo cơ chế bóng bàn. Trong nửa phản ứng đầu tiên, đơn vị galactose được chuyển từ galactinol sang enzyme, tạo thành phức hợp enzyme-galactose trung gian:
Galactinol + E ⇔ myo ‐ inositol + [E ‐ Gal] (1)
Trong nửa sau phản ứng, đơn vị galactose được chuyển sang raffinose (hoạt động STS) hoặc stachyose (hoạt động VBS):
[E ‐ Gal] + raffinose ⇔ E + stachyose (2) [E ‐ Gal] + stachyose ⇔ E + verbascose (3)
Tất cả các phản ứng đều có tính chất thuận nghịch, xảy ra tại một vị trí hoạt động duy nhất. Do đó, sự đảo ngược của một nửa phản ứng (Phương trình 2) và một nửa phản ứng (Phương trình 3) có thể được kết hợp như một chu trình xúc tác toàn bộ, tạo ra verbascose và raffinose từ stachyose làm chất nền duy nhất:
Stachyose + stachyose ⇔ raffinose + verbascose (4)
Trong một nghiên cứu về sự tích tụ của RFOs trong hai giống đậu tương cho thấy yếu tố giống cũng ảnh hưởng đến sự tích tụ đường verbascose. Cụ thể, sự tích tụ của RFOs được nghiên cứu trong hạt chín của hai dòng đậu (Pisum sativum), hạt của dòng SD1 tích lũy stachyose là chiếm ưu thế trong khi verbascose dường như không có mặt. Tuy nhiên, hạt của dòng RRRbRb có một lượng verbascose được tích lũy cùng với stachyose. Sự gia tăng verbascose trong việc phát triển hạt RRRbRb có liên quan đến hoạt động tổng hợp verbascose và phụ thuộc galactinol mà cụ thể là các mRNA liên quan đến việc mã hoá các enzyme [30].
1.2.2 Con đường sinh tổng hợp RFOs trong cây đậu tương và các enzyme tham gia
1.2.2.1 Con đường sinh tổng hợp RFOs trong cây đậu tương
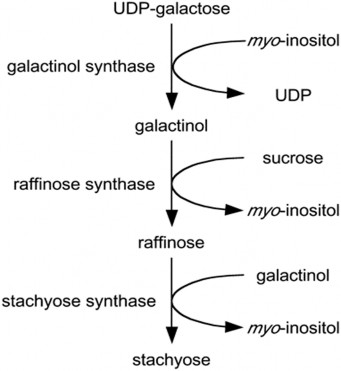
Quá trình sinh tổng hợp RFOs ở thực vật bao gồm một chuỗi các phản ứng được thực hiện để chuyển nhóm chức (nhóm hoạt tính) galactosyl từ các hợp chất nhượng nhóm chức (bao gồm: UDPD-galactose, galactinol và từ chính các hợp chất RFOs) sang các phần tử nhận nhóm chức điển hình như đường sucrose, raffinose hay các hợp chất có đồng vị cao với nó (Hình 1.4) [31]. Nhiều nghiên cứu cho thấy sự tổng hợp RFOs thường diễn ra ở lá, cụ thể là lớp trung bì của màng tế bào và các tế bào trung gian nằm ở tượng tầng libe, sau đó sẽ được vận chuyển đến các bộ khác
trong cây và tích lũy nhiều nhất ở hạt [30]. Quá trình tổng hợp RFOs ở thực vật diễn ra như sau:
Bước một (1): dưới tác động của enzyme GOLS, galactinol được hình thành từ hai thành phần ban đầu là UDP-galactose và myo-inositiol.
Bước hai (2): enzyme raffinose synthase (RS) xúc tác chuyển vị nhóm galactosyl từ galactinol sang vị trí các bon số 6 (C6) của đơn vị đường glucose trong phân tử đường sucrose, tạo lên liên kết α-1,6-galactosidic nhằm hình thành hợp chất đường ba phân tử raffinose [32].
Bước ba (3) của quá trình sinh tổng hợp RFOs: enzyme stachyose synthase xúc tác việc chuyển đơn vị galactosyl từ galactinol sang vị trí cacbon số 6 của đơn vị đường galactose trong thành phần của raffinose để hình thành hợp chất với bốn đơn vị đường đơn stachyose [33].
Ở đậu tương, quá trình sinh tổng hợp raffinose xảy ra trong hạt đang phát triển được xúc tác bởi raffinose synthase 2 (RS2), được mã hóa bởi Glyma06g18890 [14]. RS2 xúc tác phản ứng giữa sucrose và galactinol tạo thành raffinose và myo- inositol. Sự chuyển đổi tiếp theo của raffinose thành stachyose và stachyose thành verbascose tạo ra tổ hợp oligosaccharides họ raffinose, tuy nhiên hàm lượng
(1)
(2)
verbascose trong hạt đậu tương là không đáng kể [17].
(3)
Hình 1.4. Con đường sinh tổng hợp các RFOs [33].
1.2.2.2 Các enzymes tham gia con đường sinh tổng hợp
Raffinose synthase (RS)
RS là enzyme thứ hai có mặt trong quá trình tổng hợp RFOs từ galactinol và sucrose, được tinh chế lần đầu tiên từ hạt Vicia faba, kế đến là mầm lúa mì và hạt đậu tương [33]. RS có độ pH tối ưu trung tính, là chất xúc tác quá trình galactosyl hóa, hoặc nghịch chuyển sucrose từ galactinol tạo ra raffinose và myo –inositol [33].
Stachyose synthase (STS)
Đây là enzyme thứ ba trong con đường tổng hợp RFOs. Enzyme tham gia tổng hợp stachyose từ galactinol và raffinose và được đề xuất là một enzyme điều hòa tổng hợp RFOs, trên cơ sở các phát hiện cho thấy hoạt động STS trên lá tăng lên song song với việc tăng nhu cầu tiêu thụ khi đậu quả của cây dưa chuột và dưa hấu; STS lần đầu tiên được phân lập và đặc trưng từ hạt giống của Phaseo lus vulgaris và sau đó được tinh chế, chiết xuất trong hạt Vicia faba và trong lá của một số cây họ bầu bí và Ajuga. Enzyme này hoạt động tối ưu ở mức pH trung tính [35].
Verbascose Synthase
Đây là enzyme xúc tác sự hình thành DP, RFOs cao hơn còn ít được quan tâm nghiên cứu. Mặc dù sự hình thành quang hợp của [14C] -verbascose từ 14CO, đã được quan sát thấy trong lá của cây Lamium maculatum và cây bầu bí Xerosicyos danguyi, không có sự tổng hợp verbascose nào được tìm thấy trong chất chiết xuất từ lá in vitro [33].
1.2.3 Gen mã hóa cho Galactinol synthase (GOLS) trong đậu tương
GOLS là một enzyme quan trọng trong quá trình sinh tổng hợp raffinose bằng cách xúc tác sự hình thành galactinol từ myo-inositol và UDP-Galactose. Việc gây mất chức năng của gen mã hóa cho GOLS làm giảm galactinol, stachyose và tăng hàm lượng sucrose đã được nghiên cứu trên Arabidopsis. Bên cạnh đó, khi tăng cường biểu hiện của các gen mã hóa enzyme tham gia quá trình sinh tổng hợp galactinol (AtGolS1, AtGolS2, AtGolS4) trong cây Arabidopsis đã làm tăng nồng độ galactinol, raffinose và tăng khả năng chống chịu oxy hóa [36]. Tuy nhiên, các nghiên cứu về chức năng của nhóm gen này trên đậu tương còn rất hạn chế. Nghiên cứu tạo đột biến từng gen riêng biệt hoặc đồng thời các gen này sẽ là cơ sở để phân tích chức năng của
các gen mã hóa cho GOLS trên đậu tương đồng thời định hướng ứng dụng trong cải tạo và nâng cao chất lượng hạt đậu tương, tạo ra sự giảm hàm lượng của raffinose và starchyose trong hạt đậu tương.
1.2.4 Nghiên cứu thay đổi hàm lượng RFOs trên cây đậu tương
GOLS là enzyme đầu tiên tham gia trong quá trình sinh tổng hợp các RFOs. GOLS xúc tác chuyển hóa L-myo-inositiol thành galactinol, đây phản ứng đầu tiên trong quá trình sinh tổng hợp đường raffinose [31]. Sự biểu hiện quá mức của gen tổng hợp galactinol trong cây Arabidopsis biến đổi gen làm tăng nồng độ galactinol nội sinh và raffinose trong điều kiện bình thường [37].
RS xúc tác chuyển hóa đường sucrose thành đường raffinose và các hợp chất cao phân tử tiếp theo như Stachyose, Verbascose và Ajugose. Trên đậu tương, trên ba gen mã hóa cho RS được xác định, trong đó gen mã hóa RS2 (Glyma06g18890) có biểu hiện mạnh nhất trong quá trình phát triển của hạt đậu tương [14]. Do vậy, việc giảm biểu hiện của gen này sẽ tăng hàm lượng đường sucrose đồng thời nhóm đường đơn như raffinose và stachyose sẽ giảm xuống. Một số dòng đậu tương mang gen đột biến RS2 được khẳng định có chứa hàm lượng thấp raffinose trong hạt [38].
Thêm vào đó, nghiên cứu trên dòng đậu tương tạo được bằng công nghệ RNAi nhằm giảm biểu hiện của gen RS2 cũng ghi nhận sự giảm xuống của đường raffinose trong hạt đậu tương và tăng hiệu suất tiêu hóa và hấp thu năng lượng của gia cầm sử dụng loại hạt này [15]. Điều này cho thấy cơ hội và ý nghĩa quan trọng trong việc nghiên cứu quá trình sinh tổng hợp raffinose trong cải tạo chất lượng hạt đậu tương. Ngoài ra, STS là enzyme đóng vai trò quan trọng trong quá trình sinh tổng hợp stachyose. Các dòng đột biến tự nhiên cho thấy sự ức chế hoạt động của enzyme này cũng làm tăng lượng đường trong hạt và giảm đáng kể hàm lượng stachyose trong hạt
[39].
1.3 Chỉnh sửa hệ gen thông qua hệ thống CRISRP/Cas9
Năm 2011-2012, hai nhóm nghiên cứu độc lập của Tiến sĩ Emmanuelle Charpentier tại trường đại học Umea và Tiến sĩ Jennifer Doudna tại trường đại học California, Berkeley đã cùng khám phá ra cơ chế hoạt động của enzyme Cas và CRISPR. Hệ thống CRISPR/Cas được xác định là một hệ miễn dịch ở sinh vật nhân sơ có khả năng chống lại các yếu tố di truyền ngoại lai như sự xâm nhập của plasmid và thể thực khuẩn vi khuẩn; CRISPR/Cas còn được gọi là hệ thống miễn dịch di
truyền hay miễn dịch thu được Cơ chế hoạt động của enzyme Cas và CRISPR dựa trên RNA bắt cặp với trình tự của vùng đệm spacer trên DNA mới xâm nhập và giúp protein Cas (CRISPR-associated) nhận diện và thực hiện cắt đứt sợi DNA; ngoài ra phức hệ gồm RNA dẫn đường và protein Cas còn có thể cắt các RNA ngoại lai xâm nhập [41].
Tháng 1/2013, Doudna và Charpentier đã nghiên cứu đột phá cắt một mẫu DNA từ tế bào người và thay thế nó bằng một đoạn mã di truyền khác [42]. Cùng lúc, một nhóm các nhà nghiên cứu khác đến từ Đại học Harvard và Viện Broad cũng đã độc lập tuyên bố đã phát triển thành công phương pháp tương tự [43]. Đặc điểm chủ yếu của hệ thống CRISPR/Cas : (1) tạo ra điểm đứt gãy sợi kép trên DNA và (2) điểm đứt gãy này được định vị một cách chính xác nhờ đoạn crRNA định hướng. Các nhà khoa học đã ví CRISPR/Cas là “chiếc kéo phân tử” có khả năng cắt bỏ các đoạn gen không mong muốn và thay bằng đoạn gen mới theo yêu cầu [44]. Năm 2015, tạp chí Science đã bình chọn CRISPR/Cas9 là công nghệ quan trọng nhất, khởi đầu kỷ nguyên mới của công nghệ sinh học, giúp chỉnh sửa thông tin di truyền của mọi tế bào một cách nhanh chóng và chính xác [45]. Và hai nhà khoa học Doudna và Charpentier với công trình nghiên cứu về CRISPR/Cas9 đã đạt giải Nobel năm 2020.
Từ đó đến nay, hệ thống CRISPR/Cas được các nhà khoa học nghiên cứu ứng dụng trong lĩnh vực y tế, dược phẩm, nông nghiệp….Hệ thống này đang được ứng dụng trong nghiên cứu điều trị bệnh ung thư bằng cách loại bỏ các phần của bộ gen chịu trách nhiệm trực tiếp đối với sự phát triển của khối u, tiêu diệt muỗi mang virus Zika gây các bệnh sốt vàng da, điều trị mù lòa, tiêu diệt HIV [46]. Thêm vào đó, CRISPR/Cas cũng được sử dụng trong nghiên cứu các bệnh do di truyền thông qua chỉnh sửa các bộ gen của phôi thai người; điều chế các loại thuốc tốt hơn; cải thiện các thuộc tính cây nông nghiệp hay động vật trong chăn nuôi…[47].
1.3.1. Nguyên lý hoạt động của hệ thống CRISRP/Cas
Hệ thống này dựa trên cơ chế “miễn dịch” của vi khuẩn chống lại sự xâm nhiễm phân tử DNA ngoại lai từ virus hoặc DNA plasmid. Khác với hệ thống “miễn dịch” dựa trên enzyme cắt giới hạn, hệ thống CRISPR/Cas dựa trên phân tử RNA để nhận diện và phá hủy DNA ngoại lai. Để bảo vệ vi khuẩn khỏi DNA ngoại lai, vi khuẩn gắn chèn một đoạn ngắn DNA ngoại lai vào DNA bộ gen tại vùng trình tự lặp lại CRISPR. Vùng trình tự này được phiên mã và được xử lý thành các đoạn RNA