DANH MỤC CÁC KÝ HIỆU, CÁC CHỮ VIẾT TẮT
Kí hiệu Tiếng Anh Tiếng Việt
AS Acetosyringone -
A. rhizogenes Agrobacterium rhizogenes -
A. tumefaciens Agrobacterium tumefaciens -
AFLP amplified fragment length polymorphism
Sự đa hình về chiều dài các phân đoạn được khuếch đại
Biallelic Biallelic Đột biến khác nhau trên 2 alen cùng 1ocus
Có thể bạn quan tâm!
-

 Nghiên cứu ứng dụng công nghệ CRISPR/Cas9 trong tạo đột biến gen GmGOLS03, GmGOLS19 trên cây đậu tương Glycine max L. Merrill nhằm giảm lượng đường họ Raffinose trong hạt - 1
Nghiên cứu ứng dụng công nghệ CRISPR/Cas9 trong tạo đột biến gen GmGOLS03, GmGOLS19 trên cây đậu tương Glycine max L. Merrill nhằm giảm lượng đường họ Raffinose trong hạt - 1 -
 Tình Hình Sản Xuất Và Chọn Tạo Giống Đậu Tương
Tình Hình Sản Xuất Và Chọn Tạo Giống Đậu Tương -
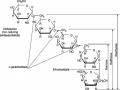 Cấu Tạo Và Mối Liên Hệ Của Các Đường Thuộc Họ Raffinose
Cấu Tạo Và Mối Liên Hệ Của Các Đường Thuộc Họ Raffinose -
![Quá Trình Hoạt Động Của Cơ Chế Crispr/cas Ở Vi Khuẩn Chống Lại Sự Xâm Nhập Của Dna Ngoại Lai [51].](https://tailieuthamkhao.com/uploads/2023/02/20/nghien-cuu-ung-dung-cong-nghe-crispr-cas9-trong-tao-dot-bien-gen-gmgols03-5-1-120x90.jpg) Quá Trình Hoạt Động Của Cơ Chế Crispr/cas Ở Vi Khuẩn Chống Lại Sự Xâm Nhập Của Dna Ngoại Lai [51].
Quá Trình Hoạt Động Của Cơ Chế Crispr/cas Ở Vi Khuẩn Chống Lại Sự Xâm Nhập Của Dna Ngoại Lai [51].
Xem toàn bộ 122 trang tài liệu này.
bp Base pair Cặp ba zơ ni tơ
Cas9 Crispr associated protein 9 Protein Cas 9

CRISPR Clustered regularly interspaced short palindromic repeat
Nhóm trình tự ngắn, phân cách đều đặn, lặp lại và đối ngẫu
DNA Deoxyribonucleic acid Axit deoxyribonucleic GOLS galactinol synthase
Gm GOLS03 Glyma.03G222000
Gm GOLS19 Glyma.19G219100
sgRNA Guide RNA Trình tự định hướng
HIV human immuno-deficiency virus
Virus gây suy giảm miễn dịch trên người
Homo Homozygous Đồng hợp tử
Hetero Heterozygous Dị hợp tử
HDR Homology directed repair Sửa chữa trực tiếp theo cơ chế
tương đồng
HPLC High-performance liquid chromatography
Sắc ký lỏng hiệu năng cao
Mr Maverick Giống đậu tương Maverick
MS Murashige and Skoog Môi trường Murashige và Skoog
NCBI National Center for Biotechnology Information
Trung tâm quốc gia về thông tin Công nghệ Sinh học Mỹ
PAM Proto-spacer adjacent motif
PCR Polymerase chain reaction Phản ứng kéo dài chuỗi nhờ
enzyme polymerase
QTL quantitative trait loci Locus liên quan tính trạng số
lượng
RNA Ribonucleic acid Axit ribonucleic
RAPD Random amplified polymorphic DNA
Ri-plasmid Root-inducing plasmid RFOs raffinose family
oligosacharides
Đa hình phân đoạn DNA được khuếch đại ngẫu nhiên
Nhóm đường họ raffinose
RFO Raffinose Đường raffinose
RS Raffinose synthase Enzyme raffinose synthase RS2 Raffinose synthase 2 Enzyme raffinose synthase 2
RFLP Restriction fragment length polymorphism
RNase III Ribonuclease III
Đa hình chiều các dài đoạn cắt bởi enzyme giới hạn
STS Stachyose synthase Enzyme sinh tổng hợp stachyose SSR Simple sequence repeat Đoạn trình tự lặp lại đơn giản
SNP Single nucleotide polymorphism
tracrRNA Trans-acting crispr RNA
Đa hình đơn nucleotide
WT Wild type Cây giống gốc hay đối chứng không chỉnh sửa gen
WL82 Williams 82 Giống đậu tương Williams 82 YEP Yeast extract peptone Môi trường Yeast extract peptone
DANH MỤC CÁC BẢNG
Trang
Bảng 2.1 Thành phần các loại môi trường trong nuôi cấy tạo rễ tơ
in vitro đậu tương. 31
Bảng 2.2 Thành phần các môi trường sử dụng trong chuyển gen cây
đậu tương. 35
Bảng 2.3 Dự kiến khả năng phát hiện các alen đột biến thông qua phản
ứng PCR sử dụng các chỉ thị phân tử 39
Bảng 3.1 Tỉ lệ mẫu sống và các mẫu biểu hiện gen chỉ thị trên môi trường chọn lọc… 50
Bảng 3.2 Kết quả chuyển gen của giống ĐT26 và Mr 55
Bảng 3.3 Danh sách các dòng đậu tương mang đột biến genGmGOLS03
và GmGOLS19……………………………………………………. 64
Bảng 3.4 Tổng hợp kiểu gen của các dòng cây T2 sau khi sàng lọc với
các chỉ thị phân tử 69
Bảng 3.5 Các thành phần khác trong hạt đậu tương 78
Bảng 3.6 Các đột biến off-target có thể có ở các cây đột biến T2 79
Bảng phụ lục 1 Bảng phụ lục 2
Trình tự các oligonucleotide và mồi sử dụng trong nghiên cứu… 101
Kết quả sàng lọc sự có mặt của gen chuyển trong các dòng đột biến… 103
DANH MỤC CÁC HÌNH VẼ VÀ ĐỒ THỊ
Trang
Hình 1.1. Biểu đồ thể hiện sản lượng cây đậu tương trên thế giới (2009- 2018) 4
Hình 1.2. Biểu đồ thể hiện sản lượng cây đậu tương ở Việt Nam (2009- 2018) 5
Hình 1.3. Cấu tạo và mối liên hệ của các đường thuộc họ Raffinose 11
Hình 1.4. Con đường sinh tổng hợp các RFOs 15
Hình 1.5. Quá trình hoạt động của cơ chế CRISPR/Cas ở vi khuẩn chống
lại sự xâm nhập của DNA ngoại lai 19
Hình 1.6. Cơ chế hoạt động của CRISPR/Cas9 trong trường hợp gây biến
đổi trình tự gen 20
Hình 1.7. Chỉnh sửa hệ gene bằng hệ thống CRISPR/Cas… 21
Hình 1.8. Chỉnh sửa bằng hệ thống CRISPR/Cas9 trên một số loại cây trồng 23
Hình 2.1. Sơ đồ minh họa nguyên lý kỹ thuật biến tính hồi tính dùng trong sàng lọc đột biến gen 34
Hình 2.2. Sơ đồ minh họa nguyên lý thiết kế các chỉ thị phân tử dùng trong sàng lọc đột biến phân ly 38
Hình 3.1. Mối quan hệ phát sinh loài giữa các protein GOLS đã được xác định trong các cây Arabidopsis, cà chua, ngô, 23 trình tự protein GOLS trên đậu tương 42
Hình 3.2. Phân tích transcriptomics cho gen GOLS trong đậu tương. 43
Hình 3.3. Cấu trúc gen GmGOLS và vị trí của các trình tự gRNA… 44
Hình 3.4. Sơ đồ thiết kế vector chỉnh sửa gen CRISPR/Cas9 để chuyển gen vào đậu tương 44
Hình 3.5. Tỉ lệ tạo rễ tơ của các giống đậu tương chuyển gen 46
Hình 3.6. Cảm ứng tạo rễ tơ trên các giống đậu tương thí nghiệm với cấu trúc pFGC/gfp trên môi trường đồng nuôi cấy 46
Hình 3.7. Cảm ứng tạo rễ tơ của các giống đậu tương thí nghiệm với cấu trúc pFGC/gfp sau 5 ngày trên môi trường cảm ứng tạo rễ 47
Hình 3.8. Quy trình cảm ứng tạo rễ tơ in vitro trên đậu tương thí nghiệm… 48 Hình 3.9. Biểu hiện gen GFP chỉ thị trên rễ tơ cây đậu
tương 51
Hình 3.10. Sản phẩm khuếch đại bằng các cặp mồi đặc hiệu được điện di trên gel agarose 1.5% trong đệm TAE 1X 52
Hình 3.11. Kết quả phân tích các đột biến ghi nhận trên các dòng rễ tơ chuyển gen 53
Hình 3.12. Minh họa quá trình chuyển gen vào đậu tương thông qua nốt lá mầm 55
Hình 3.13. Phân tích sản phẩm PCR của các dòng đậu tương đột biến gen thế hệ T0 bằng kỹ thuật biến tính - hồi tính trên gel
polyacrylamide 15% 57
Hình 3.14. Trình tự của các vùng trình tự đích trên gen GmGOLS03 (A) và gen GmGOLS19 (B) ở cây T0 58
Hình 3.15. Sự phân ly của các đột biến ở thế hệ T1 và T2 của dòng DT1.1… 60 Hình 3.16. Kết quả phân tích sự phân ly của các đột biến trên gen
GmGOLS 61
Hình 3.17. Sự phân ly của các đột biến trên gen GmGOLS ở thế hệ T2 của dòng DT1.1 62
Hình 3.18. Kết quả phân tích sản phẩm PCR của chỉ thị phân tử với các cây
đột biến T2 66
Hình 3.19. Kết quả phân tích sản phẩm PCR của chỉ thị phân tử GOLS-
seg F1 và GOLS-seg F3 với các cây đột biến T2 68
Hình 3.20 Kết quả giải trình tự vùng gen đột biến trên các cây T2 từ dòng
3.1 và 4.1 của thế hệ T0 giống Mr 70
Hình 3.21. Hình thái của các dòng đậu tương đột biến trồng trong nhà lưới.. 71 Hình 3.22. Biểu đồ thể hiện sự sinh trưởng và khối lượng hạt của các dòng
đậu tương đột biến 72
Hình 3.23. Khảo sát khả năng nảy mầm của hạt đậu tương đột biến gen 73
Hình 3.24. Tổng số cacbohydrat hòa tan và tổng số RFOs được đo bằng HPLC 75
Hình 3.25. Thành phần carbohydrate trong hạt đậu tương 76
Hình 3.26. Tỉ lệ carbohydrate dạng stachyose và sucrose trên tổng khối
lượng carbohydrate hòa tan trong hạt đậu tương 77
Hình 3.27. Tỉ lệ các thành phần khác trong hạt đậu tương 78
Hình 3.28. Phân tích trình tự off-target trong genome của các dòng đậu tương đột biến thế hệ T2 80
Hình 3.29. Xác định các dòng đột biến không mang gen chuyển 81
1. Tính cấp thiết của đề tài
MỞ ĐẦU
Đậu tương (Glycine max (L.) Merrill), thuộc họ đậu (Fabaceae) là loại cây công nghiệp có giàu dinh dưỡng và có giá trị kinh tế cao, được sử dụng làm thực phẩm cho người, thức ăn trong chăn nuôi, làm nguyên liệu cho ngành công nghiệp và xuất khẩu. Cây đậu tương có tính năng cải tạo đất trồng rất tốt. Hạt đậu tương là bộ phận có giá trị dinh dưỡng cao. Thành phần của hạt bao gồm glucid (12,5-25%), lidpid (12,5-25%) được xem là nguồn dầu thực vật chủ lực trên toàn cầu, protein (35- 50%) có giá trị dinh dưỡng tương đương với protein có nguồn gốc từ động vật và chiếm khoảng 69% lượng protein trên toàn cầu phục vụ cho ngành chăn nuôi gia súc, gia cầm. Ngoài ra, thành phần hạt còn chứa các vitamin nhóm (B, D, E…), một số các axit amin thiết yếu và các thành phần muối khoáng. Tuy nhiên, ngoài những thành phần dinh dưỡng giúp ích cho sức khỏe con người, trong hạt đậu tương lại có sự hiện diện của nhóm đường họ raffinose (Raffinose family oligosaccharide - RFOs) đây là nhóm đường khó tiêu chiếm 50% tổng lượng cacbon hòa tan trong hạt. Nhóm đường RFOs chứa liên kết α-galactosidic (1-6) chỉ có thể phân giải bởi enzyme galactosidase, tuy nhiên trong hệ tiêu hóa của người và nhóm động vật không nhai lại không chứa enzyme này. Thế nên, khi RFOs vào hệ tiêu hóa của người và nhóm động vật không nhai lại thì không được tiêu hóa ở dạ dày, đến ruột non nhóm đường này không được huyển hóa, không được hấp thu. Khi RFOs vận chuyển đến đại tràng được nhóm vi sinh vật ở đây lên men tạo ra carbon dioxide, hydro và mêtan gây ra hiện tượng đầy hơi, khó chịu, ngăn cản quá trình tiêu hóa. Chính nguyên nhân này đã làm giảm đi giá trị dinh dưỡng trong hạt đậu tương.
Những năm qua, các nhà khoa học đã áp dụng nhiều biện pháp nghiên cứu chọn tạo giống theo hướng truyền thống như lại tạo, đột biến, chuyển gen hay công nghệ RNAi nhằm giảm thiểu thành phần RFOs nâng cao giá trị dinh dưỡng của hạt đậu tương. Tuy nhiên các phương pháp này chưa thật sự mang lại hiệu quả. Gần đây, sự phát triển của công nghệ chỉnh sửa gen CRISPR/Cas9 mở ra cơ hội trong việc nghiên cứu chức năng gen ở mức độ DNA và được ứng dụng rộng rãi trong cải tạo giống cây trồng, đặc biệt trong đó có cây đậu tương. CRISPR/Cas9 với những ưu điểm như dễ dàng thiết kế cấu trúc chuyển gen, có thể tác động tới nhiều gen cùng một lúc với hiệu quả cao và đặc biệt là tạo được các dòng cây đột biến không mang
gen chuyển, công nghệ này đang được phát triển và ứng dụng rộng rãi trên nhiều loại thực vật. Đặc biệt, các nghiên cứu gần đây trên cây đậu tương cho thấy CRISPR/Cas9 đã được áp thành công để tạo các dòng đậu tương đột biến định hướng và không mang gen chuyển.
Galactinol synthase (GOLS) là enzyme quan trọng xúc tác phản ứng chuyển hóa L-myo-inositiol thành galactinol. Đây là phản ứng đầu tiên trong con đường sinh tổng hợp đường khó tiêu họ raffinose. Trên đậu tương, các nghiên cứu đã xác định được 6 gen mã hóa cho enzyme GOLS, trong đó gen Glyma.03G222000 (GmGOLS03) và Glyma.19G219100 (GmGOLS19) có biểu hiện mạnh nhất trong quá trình hạt phát triển. Việc giảm hàm lượng Galactinol thông qua đột biến định hướng trên từng gen riêng biệt hoặc đồng thời cả hai gen (GmGOLS03, GmGOLS19) được kỳ vọng sẽ tăng hàm lượng sucrose, đồng thời giảm hàm lượng đường khó tiêu RFOs có trong hạt đậu tương. Các nghiên cứu trước đây về việc điều khiển biểu hiện gen mã hóa các enzyme xúc tác trong quá sinh tổng hợp nhóm đường RFOs nhằm giảm hàm lượng nhóm đường này trong hạt đậu tương đã thu được những kết quả khả quan. Trong đó, nghiên cứu giảm biểu hiện của các gen mã hóa cho enzyme raffinose synthase và stachyose synthase đã được thực hiện thành công trên đậu tương, ghi nhận việc giảm đáng kể hàm lượng raffinose và stachyose trong hạt. Điều này là minh chứng khoa học cho việc điều khiển hoạt động của các enzyme tham gia quá trình sinh tổng hợp đường khó tiêu họ raffinose trong nâng cao chất lượng hạt đậu tương.
Xuất phát từ cơ sở khoa học và nhu cầu thực tiễn về tạo giống đậu tương có hàm lượng đường RFOs thấp, chúng tôi tiến hành thực hiện đề tài “Nghiên cứu ứng dụng công nghệ CRISPR/Cas9 trong tạo đột biến gen GmGOLS03, GmGOLS19 trên cây đậu tương (Glycine max (L.) Merrill) nhằm giảm lượng đường họ Raffinose trong hạt”
2. Mục tiêu của đề tài
Phát triển và ứng dụng hệ thống CRISPR/Cas9 để tạo đột biến định hướng trên gen mã hóa GOLS nhằm nâng cao chất lượng hạt đậu tương.
Mục tiêu cụ thể
- Thiết kế được hệ thống CRISPR/Cas9 và xây dựng thành công phương pháp kiểm tra hoạt động gây tạo đột biến gen của hệ thống này trên các giống đậu tương Việt Nam.