bình cao hơn 47,44% ở đột biến đơn (p = 0,0254) và 41,7% ở đột biến kép (p = 0,0472).
Tuy nhiên, hàm lượng raffinose của các dòng đột biến đơn và đột biến kép không khác biệt về mặt thống kê (p = 0,24). Hơn nữa, hàm lượng verbascose giảm 22,6% (ở hạt đột biến đơn) và 42,1% (ở hạt đột biến kép) so với đối chứng.
Cabonhydrates tổng số
Đường raffinose tổng số
Hình 3.24. Tổng số cacbohydrat hòa tan và tổng số RFOs được đo bằng HPLC (ĐT26 là hạt giống đối chứng; D1.1-7-2, D1.1-14-3 là hạt từ cây T2 đột biến đơn gen GmGOLS03; D1.1-5-4, D1.1-7-1 là hạt từ cây T2 đột biến hai gen GmGOLS03 GmGOLS19. Phân tích thống kê được thực hiện bằng cách sử dụng phương pháp phân tích phương sai một yếu tố (one-way ANOVA). Các chữ cái khác nhau biểu thị sự khác biệt có ý nghĩa ở mức p < 0,05. Giá trị trung bình ± SD được hiển thị cho n = 4)
Tổng hàm lượng RFOs, được đo bằng tổng của raffinose, stachyose và verbascose là 64,7 mg/g khối lượng khô ở hạt đối chứng, đã giảm 30,2% xuống 45,13 mg/g ở thể đột biến đơn và 34,1% xuống 41,95 mg/g ở đột biến kép. Trong khi đó, không tìm thấy sự thay đổi đáng kể về hàm lượng glucose hoặc fructose giữa hạt đối chứng và hạt đột biến (p > 0,05).
Hình 3.25. Thành phần carbohydrate trong hạt đậu tương
(A). Carbohydrate chiếm lượng lớn là stachyose và sucrose. (B) các carbohydrate lượng nhỏ hơn.
ĐT26 là hạt giống đối chứng; D1.1-7-2, D1.1-14-3 là hạt từ cây T2 đột biến đơn gen GmGOLS03; D1.1-5-4, D1.1-7-1 là hạt từ cây T2 đột biến hai gen GmGOLS03 GmGOLS19. Phân tích thống kê được thực hiện bằng cách sử dụng phương pháp phân tích phương sai một yếu tố (one-way ANOVA). Các chữ cái khác nhau biểu thị sự khác biệt có ý nghĩa ở mức p < 0,05.Giá trị trung bình ± SD được hiển thị cho n = 4.
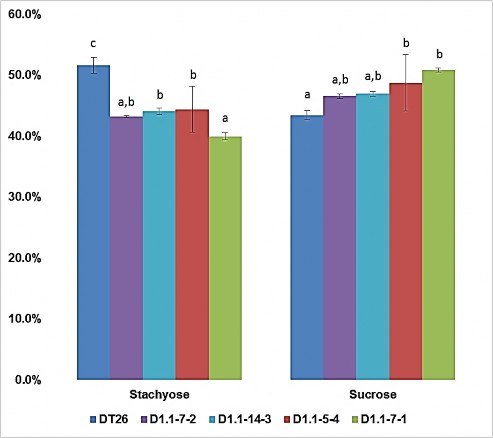
Hình 3.26. Tỉ lệ carbohydrate dạng stachyose và sucrose trên tổng khối lượng carbohydrate hòa tan trong hạt đậu tương
ĐT26 là hạt giống đối chứng; D1.1-7-2, D1.1-14-3 là hạt từ cây T2 đột biến đơn gen GmGOLS03; D1.1-5-4, D1.1-7-1 là hạt từ cây T2 đột biến hai gen
GmGOLS03 GmGOLS19. Phân tích thống kê được thực hiện bằng cách sử dụng phương pháp phân tích phương sai một yếu tố (one-way ANOVA). Các chữ cái khác nhau biểu thị sự khác biệt có ý nghĩa ở mức p < 0,05. Giá trị trung bình ± SD được hiển thị cho n = 4.
Điều đặc biệt là các dạng carbohydrate riêng lẻ được tính theo tỉ lệ phần trăm với carbohydrate hòa tan tổng số (Hình 3.26), dễ thấy là tỉ lệ tương đối của sucrose thực sự tăng lên ở cả đột biến đơn và kép so với đối chứng. Ngoài ra, tỉ lệ stachyose ở cây đột biến kép và đơn thấp hơn so với cây đối chứng.
3.5.3.2 Phân tích các thành phần khác có trong hạt của các dòng đậu tương mang đột biến
Tiến hành phân tích định lượng các thành phần khác của hạt bao gồm độ ẩm, protein, chất béo và tinh bột (Hình 3.27, Bảng 3.5).
Bảng 3.5 Các thành phần khác trong hạt đậu
Protein (%) | Chất béo (%) | Tinh bột (%) | |
WT (ĐT26) | 38,30 ± 0,49 a | 19,83 ± 1,28 a | 5,090 ± 0,84 a |
D1.1-7-2 | 40,06 ± 1,09 b | 22,88 ± 0,52 b | 5,770 ± 1,00 a |
D1.1-14-3 | 40,06 ± 0,54 b | 20,92 ± 0,69 a | 4,775 ± 0,38 a |
D1.1-5-4 | 38,28 ± 0,49 a | 20,37 ± 1,45 a | 5,509 ± 1,29 a |
D1.1-7-1 | 38,68 ± 0,53 a | 20,52 ± 0,79 a | 5,219 ± 1,19 a |
Có thể bạn quan tâm!
-
 Kiểm Tra Hoạt Động Của Cấu Trúc Chỉnh Sửa Gen Crispr/cas9 Trên Các Dòng Rễ Tơ
Kiểm Tra Hoạt Động Của Cấu Trúc Chỉnh Sửa Gen Crispr/cas9 Trên Các Dòng Rễ Tơ -
 Sàng Lọc Các Dòng Đậu Tương Chỉnh Sửa Gen Qua Các Thế Hệ T1 Và T2
Sàng Lọc Các Dòng Đậu Tương Chỉnh Sửa Gen Qua Các Thế Hệ T1 Và T2 -
 Kết Quả Phân Tích Sản Phẩm Pcr Của Chỉ Thị Phân Tử Gols-Seg F1 Và Gols-Seg F3 Với Các Cây Đột Biến T2
Kết Quả Phân Tích Sản Phẩm Pcr Của Chỉ Thị Phân Tử Gols-Seg F1 Và Gols-Seg F3 Với Các Cây Đột Biến T2 -
 Việc Lựa Chọn Trình Tự Mục Tiêu Trong Thiết Kế Cấu Trúc Chỉnh Sửa Gen Trên Đậu Tương
Việc Lựa Chọn Trình Tự Mục Tiêu Trong Thiết Kế Cấu Trúc Chỉnh Sửa Gen Trên Đậu Tương -
 Nghiên cứu ứng dụng công nghệ CRISPR/Cas9 trong tạo đột biến gen GmGOLS03, GmGOLS19 trên cây đậu tương Glycine max L. Merrill nhằm giảm lượng đường họ Raffinose trong hạt - 14
Nghiên cứu ứng dụng công nghệ CRISPR/Cas9 trong tạo đột biến gen GmGOLS03, GmGOLS19 trên cây đậu tương Glycine max L. Merrill nhằm giảm lượng đường họ Raffinose trong hạt - 14 -
 Nghiên cứu ứng dụng công nghệ CRISPR/Cas9 trong tạo đột biến gen GmGOLS03, GmGOLS19 trên cây đậu tương Glycine max L. Merrill nhằm giảm lượng đường họ Raffinose trong hạt - 15
Nghiên cứu ứng dụng công nghệ CRISPR/Cas9 trong tạo đột biến gen GmGOLS03, GmGOLS19 trên cây đậu tương Glycine max L. Merrill nhằm giảm lượng đường họ Raffinose trong hạt - 15
Xem toàn bộ 122 trang tài liệu này.
Ghi chú: Hạt WT (wild type) là giống ĐT26 không mang đột biến; D1.1-7-2, D1.1-14-3 là tên các dòng đậu tương T2 đột biến đơn gen GmGOLS03; D1.1-5-4, D1.1-7-1 là tên các dòng đậu tương T2 đột biến đồng thời hai gen GmGOLS03 và GmGOLS19 double mutants. Số liệu được phân tích thống kê phương sai ANOVA và kiểm định hậu định bằng kiểm định Tukey ở mức ý nghĩa p=0,05.
Tinh bột Độ ẩm Chất béo
Protein
Hình 3.27. Tỉ lệ các thành phần khác trong hạt đậu tương
ĐT26: hạt dòng đối chứng; DT1.1-7-2, DT1.1-14-3: hạt các dòng T2 mang đột biến đơn gen GmGOLS03; DT1.1-5-4, DT1.1-7-1: hạt các dòng T2 mang đột biến hai gen GmGOLS03 và GmGOLS19
Kết quả phân tích không có sự khác biệt đáng kể về thành phần tinh bột trong hạt giữa hạt đột biến được kiểm tra và hạt đối chứng. Tuy nhiên, hạt từ các dòng đột
biến đơn GmGOLS03 có hàm lượng protein tăng (40,06% so với 38,30% của đối chứng, p = 0,00254) và hàm lượng chất béo tăng (21,9% so với 19,8% của đối chứng, p = 0,0201), trong khi ở các dòng đột biến kép GmGOLS03 và GmGOLS19 không có sự thay đổi đáng kể nào (p = 0,581 và 0,406 lần lượt đối với chất đạm và chất béo).
Kết quả này cho thấy chức năng của GmGOLS03 và GmGOLS19 không chỉ giới hạn trong quá trình sinh tổng hợp RFOs mà nó còn có thể liên quan đến quá trình sinh tổng hợp của một số thành phần khác có trong hạt đậu tương và các nghiên cứu sâu hơn cần được thực hiện để làm rõ kết luận này.
3.6. Kiểm tra đột biến ngoài định hướng và lựa chọn dòng đột biến tiềm năng không mang gen chuyển
Để đảm bảo tính ứng dụng của các dòng đậu tương đột biến triển vọng với hàm lượng đường họ raffinose thấp trong hạt, việc quan trọng là cần khẳng định sự vắng mặt của các đột biến ngoài mục tiêu cũng như chọn được các dòng đột biến không mang gen chuyển. Sau khi đã lựa chọn được các dòng đậu tương mang đột biến ổn định dạng đồng hợp tử, tiến hành phân tích các đột biến ngoài mục tiêu và sàng lọc các dòng đột biến không mang gen chuyển.
3.6.1. Phân tích đột biến ngoài mục tiêu (off target)
Hoạt động ngoài mục tiêu (off-target) đã được báo cáo trong một số nghiên cứu sử dụng hệ thống CRISPR/Cas9 [83].
Bảng 3.6 Các đột biến off-target có thể có ở các cây đột biến T2
Trình tự | MMs | Vị trí gen | Vùng chức năng | Đột biến được xác định | |
Trình tự đích | GAGTCACACCCCTCAGT ACA | 0 | Glyma.03G38080 Glyma.19G40680 | Exon | |
Off- target có thể có | GAGTCAAGCCCCACAGT ACATGG | 3 | Glyma.07G34570 | UTR | 0 |
Trình tự đích | GCACCTTCTCCGGGCATTG C | 0 | Glyma.03G38080 Glyma.19G40680 | Exon | |
Off- target có thể có | GCACTTACTTCGGGGATT GCAGG | 4 | Glyma.09G14090 | Exon | 0 |
Ghi chú: Chữ đỏ trong trình tự thể hiện vị trí sai khác so với trình tự đích, chữ gạch chân là trình tự nhận biết PAM; MMs: số vị trí sai khác so với trình tự định hướng thiết kế.
Trong quá trình lựa chọn mục tiêu bằng chương trình trực tuyến CCTop [93], hai vị trí có khả năng gây tạo đột biến ngoài trình tự mục tiêu lần lượt nằm trong gen Glyma.07G220600 và Glyma.09G169400 đã được xác định. Cả trình tự genome của giống đối chứng William 82 và các giống thử nghiệm (ĐT26 và Mr) đều chứa các vị trí ngoài mục tiêu tiềm năng có trình tự PAM giống hệt nhau và có 3-4 nucleotide không khớp bất cứ mục tiêu nào được sử dụng (Bảng 3.6).
Chúng tôi thực hiện các phân tích ngoài mục tiêu trên các dòng đột biến T2 và T3 bằng cách giải trình tự các vùng bên ngoài mục tiêu trên hai gen Glyma.07G220600 và Glyma.09G169400 (Hình 3.28).
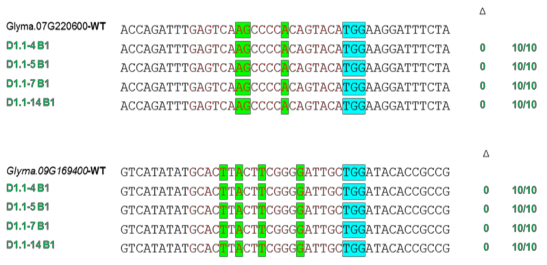
Kết quả cho thấy không phát hiện hoạt động chỉnh sửa nào tại các vị trí ngoài mục tiêu, điều này cho thấy tính đặc hiệu cao trong chỉnh sửa gen sử dụng hệ thống CRISPR/Cas9 ở đậu tương.
Hình 3.28. Phân tích trình tự off-target trong genome của các dòng đậu tương đột biến thế hệ T2
Vùng tô xanh lá thể hiện trình tự sai khác so với trình tự đích, vùng tô xanh dương thể hiện trình tự nhận biết PAM
3.6.2. Xác định các đột biến đồng hợp tử không mang gen chuyển
Tiến hành sàng lọc các gen chuyển từ thế hệ T0 đến T2 bằng phương pháp phết thuốc diệt cỏ trên lá, kết hợp phân tích PCR mẫu DNA để khẳng định sự di truyền gen bar, kết quả sàng lọc thể hiện tại Bảng 3.3 và Hình 3.29. Ở thế hệ T1, tất cả 29 mẫu cây T1 của dòng DT1.1 được sàng lọc đều cho kết quả kháng thuốc diệt cỏ, đồng thời cho kết quả dương tính khi phân tích bằng PCR với cặp mồi đặc hiệu pcoCas9 và 35SPPDK (Bảng phụ lục 1), điều này đã khẳng định sự có mặt của gen
chuyển. Bên cạnh đó, một số cây T1 có nguồn gốc từ các dòng chuyển gen M3.1 và M4.1 cho kết quả đối lập, từ đó khẳng định các cây T1 này có kiểu gen không mang gen chuyển.
Đối chứng
Hình 3.29. Xác định các dòng đột biến không mang gen chuyển
Tiếp tục sàng lọc thế hệ T2 của dòng DT1.1, chúng tôi xác định được dòng DT1.1-7-1, có kiểu gen đột biến GmGOLS kép và dòng DT1.1-7-2 mang đột biến đơn của gen GmGOLS03 đều không mang gen kháng thuốc trừ cỏ. Tất cả các cây T3 được tạo ra từ hai dòng đột biến này đều mẫn cảm với thuốc diệt cỏ. Kết quả phân tích PCR cũng cho thấy không có sự biểu hiện gen bar và gen Cas9 trong mẫu DNA của các cây T3. Điều này khẳng định, cấu trúc chuyển gen đã phân ly khỏi các dòng đột biến ở thế hệ T2 và T3.
Từ các kết quả phân tích trên, chúng tôi đã lựa chọn được các dòng đột biến tiềm năng dạng đồng hợp tử, có hàm lượng đường họ raffinose trong hạt thấp, không mang gen chuyển và không chứa các đột biến ngoài mục tiêu. Các dòng đột biến triển vọng này là nguồn nguyên liệu quan trọng cho công tác chọn tạo giống đậu tương Việt Nam theo định hướng nâng cao chất lượng hạt trong tương lai.
CHƯƠNG 4. THẢO LUẬN
So sánh với các công trình trước đây trong cùng lĩnh vực và hướng nghiên cứu, chúng tôi thấy được những phát hiện mới, những đóng góp khoa học từ kết quả nghiên cứu trong luận án này. Các thông tin này được chúng tôi thảo luận kỹ trong các phần dưới đây:
4.1 Hiệu quả chuyển gen và chỉnh sửa gen của hệ thống cảm ứng tạo rễ tơ trên các giống đậu tương Việt Nam
Việc chuyển gen bền vững và ổn định vào cây đậu tương thường sử dụng nguyên liệu là lá mầm và đỉnh sinh trưởng. Tuy nhiên, hiệu quả của phương pháp này không cao, đòi hỏi có một quy trình tối ưu và cần nhiều thời gian, chi phí để hoàn thiện một quy trình và tạo được cây đậu tương chuyển gen. Do đó, hệ thống rễ tơ được xem là phương pháp hiệu quả trong các nghiên cứu về quá trình cộng sinh, khả năng hấp thu dinh dưỡng, tương tác với tác nhân gây bệnh và đặc biệt trong nghiên cứu chức năng gen hay chỉnh sửa gen trên đậu tương.
Kết quả xây dựng hệ thống cảm ứng tạo rễ tơ và chuyển gen vào rễ tơ trên một số giống đậu tương nghiên cứu được trình bày ở mục 3.2.1. So với nghiên cứu của Chen và đồng tác giả năm 2018, lá mầm đậu tương 5 ngày tuổi bao gồm cuống lá dài 0,5 cm được sử dụng làm nguyên liệu biến nạp [81]. Tỉ lệ cảm ứng tạo rễ tơ của các giống đậu tương trong nghiên cứu này đạt 90-99%. Tuy nhiên, hiệu suất chuyển gen chỉ đạt từ 30-60%. Tương tự như vậy, khi mầm 5 ngày tuổi được sử dụng trong biến nạp gen gfp trong công bố của Keyes và đồng tác giả năm 2009, tỉ lệ cảm ứng tạo rễ tơ chỉ đạt 45%, trong đó số rễ tơ mang và biểu hiện gen chuyển chỉ đạt khoảng 55% [94].
Phương pháp cảm ứng tạo rễ tơ trong điều kiện in vitro với vật liệu là lá mầm 3 ngày tuổi của các giống đậu tương Việt Nam, chúng tôi đã thu được tỉ lệ chuyển gen cao nhất là 79,8% với giống ĐT26 khi sử dụng cấu trúc mang gen gfp. Tỉ lệ cảm ứng tạo rễ tơ cũng đạt trên 90% với giống đậu tương này. Phương pháp này đã cho kết quả biểu hiện gen trong thời gian ngắn hơn giúp giảm thiểu thời gian và chi phí cho việc kiểm tra hoạt động của các cấu trúc chuyển gen.
Hệ thống CRISPR/Cas9 đã được áp dụng thành công để tạo đột biến gen thông