CHƯƠNG 1: TỔNG QUAN TÀI LIỆU
1.1. Giới thiệu chung về nấm Purpureocillium lilacinum
1.1.1 Vị trí phân loại
Giới (Kingdom) : Nấm
Ngành (Division): Ascomycota
Lớp (Class): Sordariomycetes
Bộ (Order): Hypocreales
Có thể bạn quan tâm!
-

 Nghiên cứu phân lập nấm Purpureocillium Lilacinum để phòng trừ tuyến trùng bướu rễ Meloidogyne SP - 1
Nghiên cứu phân lập nấm Purpureocillium Lilacinum để phòng trừ tuyến trùng bướu rễ Meloidogyne SP - 1 -
![A. Nội Bào Tử Của Vi Khuẩn Pasteuria Penetrans, B Và C: Cơ Chế Tiệu Diệt Tuyến Trùng Của Vi Khuẩn Pasteuria Penetrans [43]](https://tailieuthamkhao.com/uploads/2022/05/29/nghien-cuu-phan-lap-nam-purpureocillium-lilacinum-de-phong-tru-tuyen-trung-buou-3-1-120x90.gif) A. Nội Bào Tử Của Vi Khuẩn Pasteuria Penetrans, B Và C: Cơ Chế Tiệu Diệt Tuyến Trùng Của Vi Khuẩn Pasteuria Penetrans [43]
A. Nội Bào Tử Của Vi Khuẩn Pasteuria Penetrans, B Và C: Cơ Chế Tiệu Diệt Tuyến Trùng Của Vi Khuẩn Pasteuria Penetrans [43] -
 Môi Trường Phân Lập, Giữ Giống Nấm Purpureocillium Lilacinum (Môi Trường Potato Glucose Agar-Pga )
Môi Trường Phân Lập, Giữ Giống Nấm Purpureocillium Lilacinum (Môi Trường Potato Glucose Agar-Pga ) -
 Phương Pháp Nghiên Cứu Ảnh Hưởng Của Môi Trường Nuôi Cấy Lên Sinh Khối Và Số Lượng Bào Tử Của Nấm Purpureocillium Lilacinum
Phương Pháp Nghiên Cứu Ảnh Hưởng Của Môi Trường Nuôi Cấy Lên Sinh Khối Và Số Lượng Bào Tử Của Nấm Purpureocillium Lilacinum
Xem toàn bộ 84 trang tài liệu này.
Họ (Family): Ophiocordycipitaceae
Chi (Genus) : Purpureocillium (theo Luangsa-ard, Hywel-Jones, Houbraken & Samson (2011))
Loài: Purpureocillium lilacinum (Thom) Luangsa-ard, Hou- braken, Hywel-Jones & Samson (2011)
Tên đồng nghĩa: Paecillium Luangsa-ard, Hywel-Jones & Samson (2007); Penicillium lilacinum Thom (1910); Penicillium amethystinum Wehmer (1923)
Spicaria rubidopurpurea Aoki (1941) Purpureocillium lilacinum (Thom) Samson (1974).
Purpureocillium là một chi nấm trong họ Ophiocordycipitaceae. Chi đơn loài có chứa loài duy nhất là Purpureocillium lilacinum, là loài nấm sợi hoại sinh phổ biến. Nó đựơc phân lập từ một loạt các môi trưòng sống bao gồm đất trồng và đất bỏ hoang, rừng, đồng cỏ, sa mạc, trầm tích cửa sông, bùn cặn nứơc thải và côn trùng. Nó cũng đã đựơc tìm thấy trong trứng giun tròn, tuyến trùng nang. Ngoài ra nó còn đựơc phát hiện thường xuyên trong vùng rễ của nhiều loại cây trồng. Chúng có thể phát triển ở phạm vi nhiệt độ rộng từ 8-38 oC, và nhiệt độ sinh trưỏng tối ưu trong khoảng 26-30 oC. Nó cũng có khả năng chịu
đựng pH rộng 2-10 và có thể phát triển trên nhiều loại bề mặt khác nhau.
Purpureocillium lilacinum đựơc cho là có tiềm năng sử dụng để kiểm soát sự phát triển của tuyến trùng rễ [16],[19].
1.1.2. Lịch sử nghiên cứu
Các loài đựơc mô tả bởi nhà nấm học người Mỹ Charles Thom vào năm 1910 với tên gọi Penicillium lilacinum.
Vào năm 1974 Charles Thom chuyển các loài này thành Paecilomyces. Những ấn phẩm trong thập niên 20 chỉ ra chi Paecilomyces không phải là đơn ngành mà có quan hệ mật thiết với Paecilomyces nostocoides, Isaria takamizusanensis and Nomuraea atypicola.
Chi mới Purpureocillium được tạo ra để tổ chức các đơn vị phân loại. Tên gọi chung này đề cập đến các bào tử tím được tạo ra bởi nấm này.
1.1.3. Đặc điểm sinh học
Purpureocillium lilacinum tạo thành một sợi nấm dầy đặc làm phát sinh cành bào tử phân sinh. Phần cuối của thể bình mang những bào tử được xếp thành chuỗi. Bào tử nảy mầm khi có độ ẩm và chất dinh dưỡng thích hợp.
Khuẩn lạc trong môi trường thạch Malt phát triển nhanh, đạt đuờng kính 5-7cm trong vòng 14 ngày ở 15oC, bao gồm những sợi nấm khí sinh ban đầu có màu trắng nhưng khi hình thành bào tử thì có sự thay đổi sắc thái thành màu đỏ rượu vang.
Sợi dinh dưỡng có thành bao trơn, trong suốt, kích thứơc 2.5-4 mm. Cuống sinh bào tử phát sinh từ sợi ký sinh hoặc phát sinh từ sợi nấm khí sinh. Cuống sinh bào tử dài khoảng 400-600 μm.
Thể bình gồm một phần đuôi phình to, thon dần ở phần đầu giống như một cái cổ. Bào tử trong chuỗi có nhiều hình dạng từ elip đến hình thoi. Bào tử có thành trơn đến hơi nhám. Không có bào tử chống chịu [19].
Hình 1.1: Khuẩn lạc nấm Purpureocillium lilacinum (hình bên trái), cấu tạo vi thể nấm
Purpureocillium lilacinum (hình bên phải) [36]
1.1.4. Vai trò của Purpureocillium lilacinum
a. Tác nhân kiểm soát tuyến trùng
Purpureocillium lilacinum là một loài nấm bông phổ biến với phạm vi phân bố rộng trên toàn thế giới (Samson, 1974) và đã được thử nghiệm rộng rãi nhất cho sự kiểm soát của bệnh sần rễ và u nang tuyến trùng.
Purpureocillium lilacinum sử dụng chủ yếu trong điều kiện nhiệt đới, ví dụ như, ở Philippines (Jatala, 1986) và Cộng hòa Nam Phi (Neethling, 2002). Loài này có hiệu quả chống lại các loài tuyến trùng khác nhau gây bệnh thực vật, và chủ yếu lây nhiễm trứng và con cái tuyến trùng (Amancho và Sasser năm 1995; Borisov năm 1998; Pandey và Trivedi, 1990; Silva và cộng sự, 1992;. Sosnowska năm 2001; Zaki, 1994). Wang và cộng sự. (2001).
Quan sát một số nhiễm trùng giai đoạn ấu trùng ở Đông Trung Quốc có
sự khác biệt lớn trong khả năng gây bệnh giữa các chủng Purpureocillium lilacinum (Rodriguez- Kabana et al., 1984). Ở Ba Lan, một dòng nội địa của loại nấm này loài được nghiên cứu như một tác nhân sinh học tiềm năng chống lại
tuyến trùng sần rễ trong nhà kính (Sosnowska, 2003).
Hiện nay, Purpureocillium lilacinum là loài nấm chỉ có mục đích thương mại để kiểm soát tuyến trùng, sâu bệnh ở châu Âu, và chủng nấm 251 thương mại được đăng ký để bán ở một số nước (Atkins et al., 2005). Bởi vì nó được phân loại như một microbiopesticide [23].
Cơ chế tấn công trứng tuyến trùng:
Trứơc khi xâm nhiễm vào trứng tuyến trùng, Purpureocillium lilacinum áp sát vào bề mặt trứng và trở nên sát gần với trứng. Purpureocillium lilacinum sản xuất những tế bào chuyên biệt điển hình để lây nhiễm ở khắp nơi trên bề mặt trứng của tuyến trùng. Những tế bào chuyên biệt tương tự như những mấu lồi ở phần cuối của một sợi nấm giúp áp sát vỏ trứng. Sau khi sợi nấm xâm nhập vào vỏ trứng chúng nhanh chóng phá huỷ ấu trùng bên trong sau đó sợi nấm tạo thành cuống sinh bào tử ra phía ngoài xâm nhiễm các trứng liền kề [19].
Hình 1.2: Cơ chế tấn công trứng tuyến trùng Meloidogyne spp. bởi nấm
Purpureocillium lilacinum [42]
Hình 1.3: Trứng của tuyến trùng Meloidogyne javanica bị nhấn chìm bởi sợi nấm
Purpureocillium lilacinum
b. Sản xuất một số enzym
Nhiều enzym đựơc sản xuất bởi Purpureocillium lilacinum đã đựơc nghiên cứu như: serine protease một loại enzym chống lại trứng của tuyến trùng Meloidogyne hapurpureocillium lilacinuma: protease và chitinase có thể làm suy yếu vỏ trứng của tuyến trùng [16],[19].
c. Kháng khuẩn
Purpureocillium lilacinum khi được nuôi cấy lên men chìm người ta đã chiết tách được hoạt chất sinh học có tên gọi là leucinostatins. Leucinostatins là một peptide trung tính với cấu trúc gồm: α-aminoisobutyric acid, L-leucine, β-alanine và theo sau bởi 3 acid amin bất thường (L-threo- β-hydroxy leucine, 2-amino-2-hydroxy-4-methyl-8 oxodecanoicacid và cis-4-methyl-L-proline (theo Mori et al. 1982).Leucinostatins có 6 loại đã được môt tả là A, B, C, D, E, F. Leucinostatins có khả năng kháng khuẩn, chống lại vi khuẩn Gram dương và nhiều loại nấm (theo Fukushima et al. 1983 a,b). Tuy nhiên trong cơ chế kiểm soát tuyến trùng không có sự tham gia của loại độc tố này [9].
1.1.5. Tình hình ứng dụng nấm Purpureocillium lilacinum trong phòng trừ tuyến trùng ở Việt Nam và trên thế giới
a. Các sản phẩm diệt tuyến trùng từ nấm Purpureocillium lilacinum trên thế giới
Một số chế phẩm diệt tuyến trùng được sản xuất từ nấm Purpureocillium lilacinum trên thị trường thế giới
Hình 1.4: Một số chế phẩm diệt tuyến trùng từ nấm Purpureocillium lilacinum trên thị trường thế giới [40]
b. Các sản phẩm diệt tuyến trùng từ nấm Purpureocillium lilacinum ở Việt Nam
+ Tuyến trùng: Sử dụng nấm đối kháng Paecilomyces (Palila 500 WP) liều lượng theo khuyến cáo, hoạt chất Cytokinin (Sincosin + Agrispon) 0,2%; Etobon 0,56 SL, tưới gốc nồng độ: 0,125 ml/lít nước; 3.000-5.000 lít nước/ha, Vimoca 10G rải quanh gốc (30g/gốc tưới đẫm nước).
PALILA : ( Purpureocillium lilacinum 500 triệu bào tử/g)
Sản phẩm đặc trị tuyến trùng, ngăn ngừa bênh chết nhanh chết chậm trên tiêu và nhiều loại cây trồng khác, làm khoẻ lại bộ rễ và hút dinh dưỡng tốt hơn.
Đặc biệt thử nghiệm hiệu quả trên cao su, cà phê, hồ tiêu, đã khảo nghiệm và chất lượng đuợc khẳng đinh tại Đơn Dương- Đức Trọng, Chư sê, Daksong….
12
Trên cây trồng: cải bắp ( sú) , hồ tiêu, cà phê…. [40].
1.2. Tuyến trùng sần rễ Meloidogyne spp.
Tuyến trùng sần rễ (root-knot nematodes) được coi là nhóm tuyến trùng ký sinh quan trọng nhất. Nhóm tuyến trùng này phân bố rộng khắp thế giới và ký sinh ở hầu hết các cây trồng quan trọng ở các vùng khí hậu khác nhau. Chúng gây nên giảm sản lượng thu hoạch cũng như chất lượng sản phẩm cây trồng. Hiện nay đã thống kê khoảng gần 80 loài ký sinh thuộc chi này, trong đó có 4 loài ký sinh gây hại quan trọng nhất là: M. incognita, M. arenaria, M. javanica và M. hapurpureocillium lilacinuma. Đây là các loài phân bố rộng và gây hại lớn ở các vùng nông nghiệp trên thế giới. Ngoài ra một số loài khác mặc dù cũng gây hại quan trọng nhưng chúng chỉ gây hại ở 1-2 cây trồng và phân bố hẹp [18].
1.2.1. Đặc trưng sinh học
Trứng của tuyến trùng sần rễ được con cái đẻ ra ngoài trong một bọc gelatine (còn gọi là bọc trứng) nằm trên bề mặt của sần rễ. Đôi khi các bọc trứng này cũng có thể nằm bên trong nốt sần. Sau quá trình phát triển phôi thai, trứng phát triển thành ấu trùng tuổi 1 ngay bên trong trứng. Lần lột xác thứ nhất xảy ra trong trứng và phát triển thành ấu trùng tuổi 2. Trứng nở ra ấu trùng tuổi 2 dạng cảm nhiễm (infective juvenile = IJ2) không cần có sự kích thích của rễ thực vật.
Hình 1.5: Tuyến trùng cái Meloidogyne sp. và khối trứng
http://www.tstcantho.com.vn/?mod=article&id=193§ion_id=3
Chuẩn bị xâm nhập vào rễ IJ2 tập trung dọc theo các tế bào non ngay tại phía sau vùng đỉnh rễ. IJ2 thường tấn công vào các mô phân sinh ở đỉnh rễ, nơi các rễ bên mọc ra tạo
nan điểm xâm nhập cho các IJ2 khác và làm cho bề mặt rễ bị tổn thương. Khi IJ2 tiếp xúc với bề mặt rễ chúng thường dùng kim hút châm chích và xâm nhập ngay vào trong rễ. Sự xâm nhập của chúng có thể xảy ra ở bất kỳ phía nào của rễ. Sau khi xâm nhập vào trong rễ tuyến trùng di chuyển giữa các tế bào vỏ rễ làm cho các tế bào bị tách dọc ra, sau đó tuyến trùng định vị tại vùng mô phân sinh của vỏ rễ và bắt đầu quá trình dinh dưỡng. Khi dinh dưỡng tuyến trùng cắm phần đầu vào các tế bào mô mạch của rễ, tiết men tiêu hóa làm cho quá trình sinh lý sinh hóa của mô rễ thay đổi và hình thành các điểm dinh dưỡng cho tuyến trùng. Vùng này gồm 5-6 tế bào khổng lồ (tế bào có nhiều nhân) được tạo thành trong vùng nhu mô hoặc vùng mô phloem, nơi đầu tuyến trùng. Đây là sự thích nghi chuyên hóa cao của tế bào, chúng được tạo ra và duy trì bằng tuyến trùng ký sinh. Cùng với sự hình thành tế bào khổng lồ các mô rễ xung quanh nơi tuyến trùng ký sinh cũng phình to ra tạo thành sần rễ (gall hoặc root-knot). Sần rễ thường được tạo thành trong vòng 1-2 ngày sau khi tuyến trùng xâm nhập. Kích thước của nốt sần liên quan đến cây chủ, số lượng IJ2 xâm nhập và loài tuyến trùng ký sinh [5].
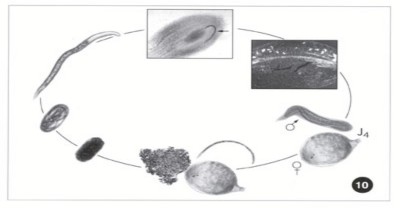
Hình 1. 6: Chu kỳ sống của tuyến trùng bướu rễ Meloidogyne sp.
http://vegetablemdonline.ppath.cornell.edu/Images/Carrots/Root-Knot/Root-KnotCycle.jpg
Bản thân tuyến trùng cảm nhiễm, sau khi xâm nhập vào rễ cũng bắt đầu một cách nhanh chóng quá trình thay đổi về hình thái: cơ thể chúng phình ra và các nội quan cũng dần được phát triển. Quá trình phát triển của tuyến trùng trong rễ từ IJ2 trải qua 3 lần lột xác và đạt đến trưởng thành. Lần lột xác cuối cùng là sự biến thái thật sự đối với con đực, từ dạng cuộn gấp khúc trong IJ4 chúng được nở ra và có dạng hình giun, trong khi đó con cái có dạng hình tròn như quả lê hay quả chanh. Tuyến trùng Meloidogyne spp. sinh sản bằng 2 cách: một vài loài sinh sản hữu tính-giao phối bắt buộc (amphimixis) và phần lớn các loài
sinh sản lưỡng tính (parthenogenessis) không cần con đực. Đối với các loài hữu tính thì con đực cặp đôi ngay với con cái sau lần lột xác cuối cùng.
Tuyến trùng sần rễ có quan hệ mật thiết với các điều kiện môi trường trong đó cây chủ, nhiệt độ và các yếu tố sinh thái đất như độ ẩm, cấu trúc đất, độ thoáng khí, độ kiềm... Có thể phân biệt 2 nhóm sinh thái liên quan đến nhiệt độ là nhóm ưa nóng (các loài điển hình như M. incognita, M. javanica, M. exigua) và nhóm ưa lạnh (các loài điển hình như M. hapla, M. chitwooodi và có thể cả M. naasi) liên quan đến pha chuyển hóa lipid của tuyến trùng xảy ra ở 10 °C. Tác hại do tuyến trùng gây ra đối với cây trồng thường có liên quan đến loại đất kiềm, là môi trường tạo ra các sốc bất lợi (stress) cho thực vật.
1.2.2. Các loài quan trọng
M. incognita: là loài phổ biến nhất, ký sinh gây hại trên nhiều cây trồng khác nhau và phân bố trên một vùng địa lý rộng từ 40 vĩ độ bắc đến 33 vĩ độ nam trên phạm vi toàn thế giới. Đây cũng là loài ký sinh gây hại phổ biến nhất trên cây trồng Việt Nam, trong đó chúng ký sinh gây hại phổ biến nhất ở: tiêu, cà phê, cà chua, bí đỏ, đu đủ, các cây họ Cà, họ Đậu, chuối..
M. javanica: là loài phổ biến thứ 2 sau loài trên và có dải phân bố tương tự. Đây là loài có khả năng chịu đựng qua mùa khô hạn trong thời gian 3-6 tháng. Ở Việt Nam, loài này ký sinh tương đối phổ biến sau loài M. incognita, gây hại chính cho các cây đậu phộng, chuối.
M. arenaria: là loài phổ biến thứ 3 sau, phân bố khắp thế giới như các loài M. incognita và
M. javanica. Đây cũng là loài ký sinh gây hại tương đối phổ biến ở Việt Nam trên các cây họ đậu.
M. graminicola: ký sinh gây hại chính cho lúa cạn ở vùng nhiệt đới và cận nhiệt đới (Đông Nam Á, Nam Phi, Mỹ). Ở ta loài này ký sinh tương đối phổ biến trên lúa cạn (giai đoạn lúa non, khi chưa ngập nước) ở đồng bằng sông Cửu Long [18].
1.2.3. Cơ sở phòng trừ tuyến trùng
Mục tiêu phòng trừ là: giảm mật độ quần thể tuyến trùng ban đầu và giảm số cây trồng bị nhiễm tuyến trùng.