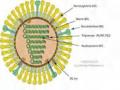
Bảng 1.1. Các phân đoạn gen của vi rút cúm A
Chiều dài đoạn gen (Nucleotide) | Protein được mã hóa | Chiều dài protein được mã hóa (Axit amin) | Chức năng | |
1 | 2341 | PB2 | 759 | Polymerase khởi đầu phiên mã |
2 | 2341 | PB1 | 757 | Kéo dài phiên mã, hoạt hóa enzyme nội bào |
PB1-F2 | 87 | Thúc đẩy quá trình chết của tế bào chủ | ||
3 | 2233 | PA | 716 | Kéo dài phiên mã |
4 | 1778 | HA | 550 | Ngưng kết hồng cầu, bám gắn xâm nhập vào tế bào chủ |
5 | 1565 | NP | 498 | Bám gắn với RNA, điều khiển quá trình nhân lên của vi rút. |
6 | 1413 | NA | 454 | Giải phóng vi rút khỏi tế bảo chủ |
7 | 1027 | M1 | 252 | Protein nền: Thành phần chính của vi rút. |
M2 | 97 | Kênh ion, cởi áo và lắp ráp thành phần vi rút | ||
8 | 890 | NS1 | 230 | Protein đối kháng interferon, điều khiển dấu ấn của vi rút trên tế bào chủ |
NS2 | 121 | Chuyển RNA vi rút từ nhân ra tế bào chất (trong tế bào chủ) |
Có thể bạn quan tâm!
-

 Tính kháng thuốc oseltamivir của virut cúm A lưu hành tại miền Bắc Việt Nam, 2001 – 2012 - 1
Tính kháng thuốc oseltamivir của virut cúm A lưu hành tại miền Bắc Việt Nam, 2001 – 2012 - 1 -
 Tính kháng thuốc oseltamivir của virut cúm A lưu hành tại miền Bắc Việt Nam, 2001 – 2012 - 2
Tính kháng thuốc oseltamivir của virut cúm A lưu hành tại miền Bắc Việt Nam, 2001 – 2012 - 2 -
 Sự Trao Đổi Và Tích Hợp Trong Hệ Gen Của Vi Rút Cúm A
Sự Trao Đổi Và Tích Hợp Trong Hệ Gen Của Vi Rút Cúm A -
 Tình Hình Kháng Thuốc Của Vi Rút Cúm A Với Thuốc Kháng Vi Rút
Tình Hình Kháng Thuốc Của Vi Rút Cúm A Với Thuốc Kháng Vi Rút -
 Kỹ Thuật Xác Định Mức Độ Kháng Oseltamivir Dựa Trên Hoạt Động Của Neuraminidase
Kỹ Thuật Xác Định Mức Độ Kháng Oseltamivir Dựa Trên Hoạt Động Của Neuraminidase
Xem toàn bộ 147 trang tài liệu này.
Trên bề mặt tế bào biểu mô đường hô hấp của người, HA gắn vào thụ thể α2,6 axit sialic; đối với gia cầm, HA gắn vào thụ thể α2,3 axit sialic; trên tế bào của lợn có mang cả hai loại thụ thể α2,3 axit sialic và α2,6 axit sialic. Ngoài ra, HA gắn vào thụ thể trên bề mặt tế bào hồng cầu người nhóm máu O, hồng cầu chuột lang, hồng cầu ngỗng, hoặc gà tây gây nên hiện tượng ngưng kết hồng cầu [118]. Năm vị trí quyết định kháng nguyên khác cũng đã được xác định trên protein HA1, tương tác với kháng thể của vật chủ. Các thay đổi có liên quan đến thay đổi đặc tính kháng nguyên xảy ra chủ yếu trên phần HA1, do vậy, HA1 được sử dụng để tìm hiểu đặc tính kháng nguyên và tiến hóa của vi rút.
Neuraminidase (NA): Protein bề mặt thứ hai có chức năng là một enzyme, neuraminidase, được mã hóa bởi phân đoạn gen thứ 6. Neuraminidase sau khi được phiên mã, protein có dạng hình nấm bao gồm 4 chuỗi polypeptide. NA xúc tác cho quá trình cắt đứt mối liên kết α2,3 hoặc α2,6 ketosidic phá hủy thụ thể HA trên bề mặt tế bào, tạo điều kiện cho sự giải phóng vi rút mới ra khỏi tế bào chủ. Việc loại bỏ phần axit sialic khỏi phần HA mới được tổng hợp còn có tác dụng tránh sự tự ngưng kết giữa các hạt vi rút mới sau khi được giải phóng ra khỏi tế bào, đảm bảo vi rút mới sau khi được giải phóng có khả năng gây nhiễm. Neuraminidase cũng đóng vai trò quan trọng trong khả năng gây bệnh của vi rút cúm. Sự hoạt động của neuraminidase làm tăng hoạt động của các cytokine, thúc đẩy nhanh quá trình chết của tế bào chủ. Ngoài ra, các nhà khoa học đã chứng minh được vi khuẩn gây nhiễm trùng thứ phát như viêm phổi đặc biệt nguy hiểm cho các bệnh nhân có bệnh mạn tính, trẻ nhỏ, người già và phụ nữ có thai, có quan hệ chặt chẽ với những hoạt động của neuraminidase như nhiễm Streptococcus pneumonia, có thể làm quá trình viêm nhiễm đường hô hấp trầm trọng hơn [122].
Vi rút cúm A được chia thành các phân típ dựa trên hai kháng nguyên bề mặt heamagglutinin (HA) và neuraminidase (NA) của vi rút. Sử dụng HA để xác định phân nhóm của vi rút cúm A, các nhà khoa học đã xác định được 17 loại hemagglutinin khác nhau, trong đó H17 được xác định từ vi rút phân lập từ dơi
[105]. Các phân típ NA chủ yếu gây bệnh trên người là N1 (H1N1, H5N1, H1N1
đại dịch), N2 (H3N2, H9N2) và gần đây là N9 (H7N9).
Dựa vào sự khác nhau về axit amin tại vùng đầu phía ngoài, neuraminidase của vi rút cúm được chia làm 2 nhóm riêng biệt [109]:
Nhóm 1 gồm N1, N4, N5 và N8
Nhóm 2 gồm N2, N3, N6, N7 và N9.
Sự thay đổi axit amin tại vị trí 147-152 đã tạo nên vùng khác biệt về hình khối nằm ngay cạnh vị trí hoạt động của N1 mà không xảy ra trên N2. Vùng khác biệt này có thể được coi là vùng đích của thuốc kháng vi rút trong tương lai.
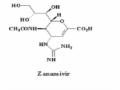
Thuốc kháng vi rút oseltaminvir, zanamivir gắn vào NA, ức chế khả năng giải phóng của vi rút cúm dựa trên hiểu biết về hoạt động của neuraminidase, xúc tác quá trình cắt đứt mối liên kết α2,3 hoặc α2,6 ketosidic giữa axit sialic và thụ thể HA. Tuy nhiên, các đột biến về axit amin trên phân đoạn gen mã hóa NA tại các vị trí bám gắn với thuốc là nguyên nhân của sự kháng thuốc của vi rút, cụ thể với N1 tại vị trí I117V, I222K, S246N, H275Y và N294S; với N2 tại các vị trí E119V, Q136K, R292K và N294S.
Protein màng (M-matrix): Đoạn RNA số 7 được coi là gen có tính bảo tồn cao, mã hóa cho hai protein M1 và M2. Protein M1 được mã hóa từ nucleotide vị tí 26 đến 784, protein M2 được tổng hợp qua quá trình ghép nối từ nucleotide vị trí 26- 51 và từ 740-1007 [19, 65]. Protein M1 là protein nền (matrix) nằm ngay dưới lớp vỏ của vi rút, làm cầu nối giữa các thành phần bên trong và các protein bề mặt của vi rút, có vai trò quan trọng trong việc lắp ráp các thành phần để tạo nên hạt vi rút, tạo điều kiện cho vi rút nảy chồi. M2 là một protein xuyên màng, phần ngoài tương tác với hệ miễn dịch của vật chủ [54] và phần xuyên màng hoạt động như một kênh ion có trách nhiệm bơm ion H+ từ nội bào vào trong vi rút, phá vỡ mối liên kết protein-protein trên protein nền M1 với RNP, giải phóng các RNP của vi rút vào trong nội bào của tế bào cảm nhiễm [3] [22, 56, 57].
Amantadine gắn vào kênh ion xuyên màng, ngăn chăn sự “cởi áo” này. Tuy nhiên, những thay đổi axit amin tại vị trí bám gắn 26, 27, 30, 31 và 34 trên kênh ion với amantadine đã tạo ra sự kháng thuốc của vi rút (vị trí gắn kết V27A Valin chuyển sang Alanin, A30T Alanin sang Threonin, S31N Serin sang Asparagin và G34E Glycin sang Axit Glutamic), trong đó vị trí 31 chuyển từ Serin sang Asparagin là vị trí có tần suất xuất hiện nhiều [47, 52]. Hiện tượng kháng amantadine xảy ra phổ biến trên gen M của vi rút cúm A/H3N2, ít gặp ở các chủng cúm A/H1N1, tuy nhiên khi chủng cúm A/H1N1pdm09 xuất hiện thì hầu hết gen M, do hiện tượng trao đổi và tích hợp gen, mang gen M của chủng vi rút cúm lợn Á Âu có đột biến kháng thuốc amantadine tại vị trí 31 (S31N) [18, 53].
Các polymerase PB1, PB2 và PA: Tồn tại trong hệ gen của vi rút cúm có các phân đoạn gen 1, 2 và 3 chịu trách nhiệm tạo ra các RNA-RNA polymerase. Phức hợp PA-PB1-PB2 ở trong nhân tế bào cảm nhiễm có nhiệm vụ khởi đầu và kích hoạt quá trình tổng hợp mRNA của vi rút [15] (hình 1.3 và hình 1.4). Trong quá trình hoạt động của vi rút cúm, đột biến xảy ra trên phân đoạn gen mã hóa PB2 có liên quan đến khả năng thích nghi của vi rút [56, 99], đặc biệt với vi rút cúm gia cầm H5N1. Đột biến E627K (Glutamin chuyển thành Lysin) trên PB2 của vi rút cúm gia cầm làm tăng khả năng thích nghi với điều kiện sống của vi rút trên vật chủ
mới - người (vi rút tồn tại và nhân lên trên gia cầm tại nhiết độ tối ưu là 37oC, khi
mang gen đột biến, vi rút có khả năng thích nghi và phát triển được trên tế bào đường hô hấp trên của người – với nhiệt độ tối ưu là 33oC) [99]. Đột biến này đã được xác định trên gen PB2 của chủng cúm gia cầm mới xuất hiện A/H7N9 [62, 63]. Một protein có kích thước nhỏ PB1-F2 (87 axit amin) có chức năng thúc đẩy quá trình chết của tế bào chủ mà không tìm thấy dạng tương tự trong hệ gen của vi rút cúm B và C [65, 122].
Nucleoprotein (NP): NP được mã hóa bởi phân đoạn thứ 5 trong bộ gen của vi rút cúm có tính bảo tồn cao đóng vai trò quan trọng trong quá trình nhân lên, tạo thành hạt vi rút mới [43]. Chức năng chủ yếu của NP là tham gia vào quá trình tổng hợp RNA của vi rút và quá trình tạo thành hạt vi rút mới. Ngoài ra, NP tương tác
với và các protein PB1 và PB2 nhưng không xảy ra với protein PA [85]. Các enzyme này khi hoạt hóa sẽ tham gia vào quá trình nhân lên và phiên mã RNA của vi rút cúm đồng thời hoạt động như một nuclease nội bào có liên hệ với ribonucleoprotein (RNP).
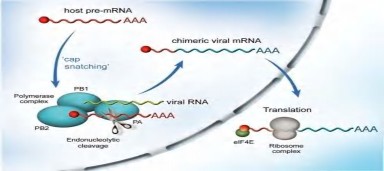
Hình 1.3. Sự tham gia của các polymerase vào quá trình sao mã của vi rút cúm Nguồn: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2937865/

Hình 1.4. Cấu trúc ribonucleoprotein của vi rút cúm Nguồn: Paul Digard, Dept. Pathology, University of Cambridge.
Protein không cấu trúc (NS1-NS2): NS1 và NS2 được mã hóa bởi đoạn RNA số 8. Phần NS1 có chức năng cản trở sự di chuyển của mRNA tế bào từ nhân ra tế bào chất tạo điều kiện cho RNA của vi rút được phiên mã ở ribosome của tế bào chủ. Ngoài ra, NS1 còn được cho là tham gia vào ức chế quá trình sản xuất interferon, chất có khả năng làm giảm sự nhân lên của vi rút cúm, của tế bào chủ. Phần NS2 của protein không cấu trúc này được tổng hợp sau NS1 và được biết với chức năng tạo thuận lợi cho việc vận chuyển các RNP mới được tổng hợp từ nhân ra tế bào chất tăng cường cho quá trình nhân lên của vi rút [43, 118, 122].
1.1.2. Cơ chế nhân lên của vi rút cúm A
Sự bám dính của vi rút
Vi rút cúm bám vào tế bào biểu mô đường hô hấp bằng cách dùng heamagglutinin gắn vào phần axit sialic của glucoprotein và glucolipid trên bề mặt tế bào. Tùy vào tế bào chủ là tế bào biểu mô đường hô hấp của người hay gia cầm mà vi rút cúm có thể sử dụng phần α2,3 hay α2,6 axit sialic trong protein HA để bám dính vào thụ cảm thể của tế bào chủ. Ngay khi quá trình bám dính hoàn thành, vi rút tiếp tục ngay quá trình thâm nhập vào tế bào chủ.
Sự thâm nhập của vi rút
Vi rút tiến vào tế bào chủ bằng quá trình thực bào, toàn bộ vi rút được bao bọc bởi màng tế bào chủ và đưa vào trong tế bào chất của tế bào chủ qua thể thực bào trong điều kiện pH thấp.
Sự cởi áo của vi rút
Sự hoạt động của protein M2 chấm dứt tình trạng pH thấp trong thể thực bào, đồng thời làm tăng cường sự hòa màng từng phần của protein HA trên màng sinh chất với màng của thể thực bào. Dòng ion từ nội bào tràn vào hạt vi rút dẫn đến sự mất liên kết giữa các protein của vi rút, khiến cho các RNP vi rút thoát khỏi sự kiểm soát của vỏ vi rút (vi rút cởi áo). Khi quá trình hòa màng đã hoàn thành, các RNP được giải phóng vào tế bào chất của tế bào chủ.
Sự tổng hợp RNA, các protein và sự giải phóng của vi rút
Khác với các vi rút mang RNA sợi đơn âm, RNA của vi rút cúm được tổng hợp và nhân lên tại nhân tế bào. Các RNP được chuyển vào nhân tế bào, phức hợp polymerase gắn vào RNA của vi rút, đồng thời cắt mRNA của tế bào chủ bằng hoạt động của nuclease nội bào. Nuclease nội bào của vi rút cắt đầu 5’ đã được gắn mũ và metyl hóa của mRNA của tế bào chủ với độ dài khoảng 13 đến 15 ba-zơ và dùng đoạn này làm mồi để tổng hợp mRNA của vi rút. Enzyme RNA polymerase của vi
rút thực hiện quá trình tổng hợp sợi mRNA bổ sung với RNA đơn âm. Phần intron (vùng câm) của mRNA vi rút bị cắt bởi các enzyme của tế bào để các protein M1 hay NS1 được tổng hợp mà không cần thêm một sự phân cắt nào. Một số protein mới được tổng hợp quay trở lại nhân tế bào gắn vào RNA để hình thành RNP. Các protein còn lại di chuyển tới lưới nội bào và thể Golgi thực hiện quá trình glucosyl hóa sau đó tiếp tục tới màng tế bào và gắn vào lớp lipid kép của màng. Khi quá trình gắn của các protein vào màng hoàn chỉnh, các RNP và M1 liên kết các thành phần lại để tạo nên hạt vi rút. Cuối cùng, hạt vi rút nảy chồi ra phía ngoài màng tế bào chủ rồi được tách ra khỏi tế bào nhờ hoạt động của neuraminidase (Hình 1.5).
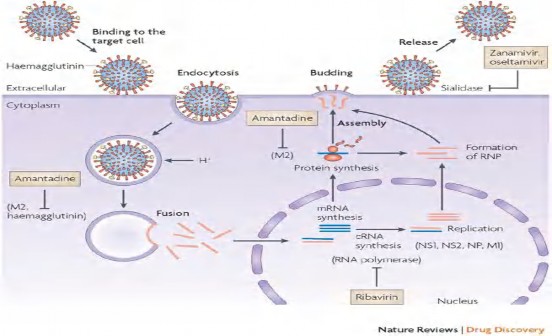
Hình 1.5. Cơ chế nhân lên của vi rút và cơ chế tác dụng của thuốc kháng vi rút Nguồn: Nature Review – Drug Discovery
1.1.3. Thay đổi nhỏ và thay đổi lớn trong hệ gen của vi rút cúm A
Những thay đổi nhỏ trên gen
Polymerase của vi rút cúm và các vi rút mang RNA nói chung không có chức năng sửa chữa sai sót trong quá trình kéo dài chuỗi sau mỗi lần nhân lên (khoảng một lỗi sai sau mỗi lần nhân lên) và đã tạo ra các biến đổi trong RNA thế hệ mới. Những biến đổi này xuất hiện một cách tự nhiên trên các phân đoạn gen của vi rút cúm nhưng chỉ làm xuất hiện những thay đổi nhỏ về đặc tính kháng nguyên của vi rút. Những biến đổi nhỏ trên phân đoạn gen mã hóa HA và NA thường được quan tâm hơn bởi những tác động của chúng lên tính lây nhiễm của vi rút cúm thế hệ mới trong quy trình gây dịch tại các năm tiếp theo, ước tính khoảng một phần trăm khác biệt trong số axit amin của HA và NA xuất hiện trong mỗi mùa dịch [6, 43, 70]. Những biến đổi nhỏ này xảy ra còn liên quan đến sự né tránh miễn dịch của vi rút cúm với kháng thể đã được tạo ra bởi những lần nhiễm trước đó, vì vậy, các vi rút mang những biến đổi này có thể là nguyên nhân gây nên những vụ dịch cúm hàng năm. Những thay đổi nhỏ xảy ra trên protein bề mặt HA và NA được ghi nhận thường xuyên nhưng lại hiếm khi với các nucleoprotein [52].
*Những thay đổi nhỏ trên phân đoạn gen mã hóa HA
Protein HA đóng vai trò quan trọng trong biểu hiện kháng nguyên của vi rút cúm. Vùng đầu chỏm của protein HA mang 5 vùng quyết định kháng nguyên, mỗi sự thay đổi trong những vùng này cũng có ảnh hưởng nhất định đến đặc tính kháng nguyên của vi rút [51, 94]. Protein HA của vi rút A/H3N2 có sự thay đổi sau mỗi 3 năm. Tuy nhiên, đáp ứng miễn dịch chéo giữa vi rút H3N2 lưu hành nổi trội với vi rút H3N2 lưu hành trước đó vẫn xảy ra [97]. Điều này chứng tỏ sự thay đổi nhỏ trên phân đoạn gen mã hóa HA chưa tạo ra được sự né tránh miễn dịch toàn bộ của vi rút cúm cho dù tần suất thay đổi nhỏ trên phân đoạn gen mã hóa HA của vi rút A/H3N2 diễn ra nhanh (3 năm), những chủng H3N2 mới vẫn chịu ảnh hưởng của miễn dịch tồn lưu được tạo ra bởi vi rút lưu hành những năm trước đó.