hoạt hóa hệ thống bổ thể theo con đường cổ điển. Hệ thống bổ thể tạo ra các hóa chất trung gian của phản ứng viêm và thúc đẩy quá trình thực bào cũng như ly giải tế bào đích. Quá trình này được bắt đầu với sự liên kết của một protein bổ thể có tên là C1q vào đoạn Fc của phân tử kháng thể IgG hoặc IgM trong phức hợp kháng nguyên – kháng thể. FcR và vị trí kết hợp bổ thể của kháng thể đều nằm ở khu vực C của chuỗi nặng của tất cả các isotyp.
Tất cả các hiệu quả chức năng của kháng thể đều chỉ được thực hiện khi kháng thể đã liên kết với kháng nguyên chứ không phải ở dạng Ig tự do. Nguyên nhân là cần phải có ít nhất 2 đoạn Fc nằm kế cận nhau để có thể tác động vào các hệ thống hiệu quả như các protein bổ thể và FcR của âaûi thực bào. Yêu cầu phải có những phân tử liền kề này đảm bảo cho chức năng hiệu quả được thực hiện một cách đặc hiệu đến đúng kháng nguyên đích và loại bỏ chúng và cũng là nhằm làm cho các kháng thể lưu động tự do trong máu không tác động thừa vào hệ thống hiệu quả.
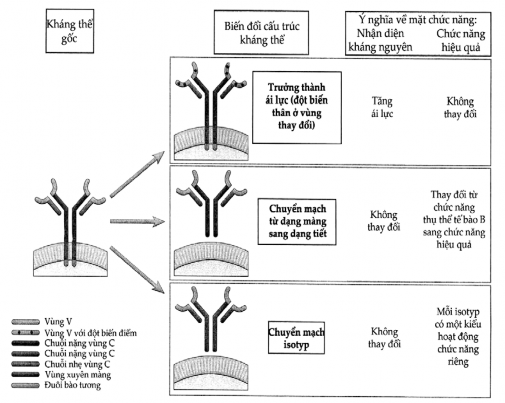
Hình 3.3. Các thay đổi của cấu trúc kháng thể trong quá trình đáp ứng miễn dịch dịch thể
Trong quá trình trưởng thành ái lực, đột biến có thể xảy ra ở vùng V dẫn đến thay đổi tính đặc hiệu trong khi chức năng hiệu quả phụ thuộc vùng C không thay đổi gì. Tế bào B hoạt hóa có thể chuyển các phân tử Ig màng sang Ig tiết. Ig tiết có thể có đột biến gen hoặc không. Trong quá trình chuyển mạch isotyp, có biến đổi ở vùng C trong khi vùng V không có gì thay đổi. Việc chuyển mạch isotyp có thể gặp ở cả Ig màng lẫn Ig tiết.
Sự thay đổi của isotyp kháng thể trong quá trình đáp ứng dịch thể sẽ
ảnh hưởng việc đáp ứng miễn dịch hoạt động loại bỏ kháng nguyên nơi nào và ở đâu. Sau khi được kháng nguyên kích thích, một clôn tế bào B sẽ sản xuất kháng thể với các isotyp khác nhau nhưng có cùng lĩnh vực V tức laì đặc hiệu cùng 1 kháng nguyên. Các tế bào B chưa tiếp xúc kháng nguyên sẽ đồng thời sản xuất IgM và IgD có tác dụng như là thụ thể bề mặt đối với kháng nguyên. Khi tế bào B tiếp xúc với một vi khuẩn chẳng hạn thì trong tế bào sẽ xảy ra một quá trình gọi là “chuyển mạch isotyp” (isotype switching) trong đó typ của vùng CH do tế bào B sản xuất bị chuyển đổi nhưng còn vùng V và tính đặc hiệu thì không thay đổi (Hình 3.3). Vì quá trình chuyển mạch isotyp mà các tế bào tiền thân của tế bào B sản xuất IgD và IgM sản xuất được các isotyp khác có khả năng tiêu diệt kháng nguyên tốt hơn. Ví dụ, kháng thể chống vi khuẩn và virus chủ yếu là typ IgG là typ có xu hướng thúc đẩy quá trình thực bào vi khuẩn, còn typ chuyên đối phó với giun tròn thì chủ yếu là IgE là typ có khả năng phá hủy ký sinh trùng này.
Có thể bạn quan tâm!
-
 Sự Chia Tách Các Vùng Của Tế Bào B Và T Trong Hạch
Sự Chia Tách Các Vùng Của Tế Bào B Và T Trong Hạch -
 Tái Tuần Hoàn Lymphô Bào Nguyên Vẹn Qua Các Cơ Quan Ngoại Biên
Tái Tuần Hoàn Lymphô Bào Nguyên Vẹn Qua Các Cơ Quan Ngoại Biên -
 Đặc Điểm Cấu Trúc Của Vùng Biến Đổi Và Mối Liên Quan Với Sự Liên Kết Kháng Nguyên
Đặc Điểm Cấu Trúc Của Vùng Biến Đổi Và Mối Liên Quan Với Sự Liên Kết Kháng Nguyên -
 Sơ Đồ Minh Họa Cấu Trúc Các Phân Tử Mhc Lớp I Và Ii
Sơ Đồ Minh Họa Cấu Trúc Các Phân Tử Mhc Lớp I Và Ii -
 Tính Chất Của Tế Bào Lymphô Nguyên Vẹn, Hiệu Quả Và Nhớ
Tính Chất Của Tế Bào Lymphô Nguyên Vẹn, Hiệu Quả Và Nhớ -
 Cố Định Và Hoạt Hóa C4 Và C2 Bởi Phức Hợp C1Qrs
Cố Định Và Hoạt Hóa C4 Và C2 Bởi Phức Hợp C1Qrs
Xem toàn bộ 103 trang tài liệu này.
Vùng C trên chuỗi nặng của kháng thể cũng quyết định vị trí phân bố của phân tử kháng thể ở các mô. IgA thường là isotyp duy nhất có thể được bài tiết qua lớp thượng bì của niêm mạc và do đó IgA là lớp kháng thể chủ yếu của các dịch tiết kể cả sữa. Trẻ sơ sinh được bảo vệ khỏi các bệnh nhiễm trùng nhờ kháng thể IgG được truyền từ mẹ sang trong quá trình mang thai và trong giai đoạn sớm của thời kỳ bú mẹ. Sự truyền này được thực hiện qua trung gian của một thụ thể Fc đặc biệt có trong bánh nhau (giúp kháng thể đi vào tuần hoàn phôi) và trong đường tiêu hóa của trẻ sơ sinh (giúp hấp thụ kháng thể từ sữa mẹ).
Chương 4
KHÁNG NGUYÊN
4.1. Các tính chất của kháng nguyên
Đáp ứng tạo kháng nguyên như ta đã biết sẽ xảy ra khi có một “vật lạ” đột nhập vào cơ thể và tiếp xúc với hệ thống miễn dịch. Vật lạ đó được gọi là chất gây kháng thể (antibody generator) hay kháng nguyên (antigen). Tuy nhiên, không phải vật lạ nào vào cơ thể cũng có tính chất kháng nguyên. Kháng nguyên có hai tính chất sau: (1) kích thích được cơ thể tạo ra đáp ứng miễn dịch, tính chất này gọi là tính sinh miễn dịch, và (2) có khả năng kết hợp đặc hiệu với kháng thể tương ứng, tính chất này là tính đặc hiệu.
4.1.1. Tính sinh miễn dịch
Tính sinh miễn dịch của một kháng nguyên phụ thuộc vào các yếu tố sau:
(1) Tính lạ của kháng nguyên: Kháng nguyên càng lạ bao nhiêu thì khả năng kích thích tạo kháng thể càng mạnh bấy nhiêu. Tuy nhiên, cần lưu ý rằng, trong một số trường hợp bệnh lý thì thành phần của chính bản thân cơ thể cũng có thể gây ra đáp ứng kháng thể chống lại nó, ta gọi những thành phần này là tự kháng nguyên.
(2) Cấu trúc hóa học của kháng nguyên: Các kháng nguyên thuộc loại protein và polysaccharid có tính sinh miễn dịch cao. Kháng nguyên càng phức tạp về cấu trúc hóa học bao nhiêu thì tính sinh miễn dịch càng mạnh bấy nhiêu. Trên cấu trúc đó có những cấu tạo chịu trách nhiệm chính trong việc kích thích tạo kháng thể, đó là các quyết định kháng nguyên hay epitop.
(3) Cách gây miễn dịch và liều kháng nguyên: Hầu hết các kháng nguyên hữu hình (vi khuẩn, hồng cầu, các polymer lớn,…) khi đưa vào cơ thể bằng đường tĩnh mạch đều dễ dàng gây đáp ứng tạo kháng thể. Trong khi đó, có một số phân tử cần phải kèm thêm một chất hỗ trợ khác mới gây được đáp ứng tốt, ta gọi chất hỗ trợ đó là tá chất adjuvant. Loại tá chất thường dùng là tá chất Freund, đó là một hỗn dịch vi khuẩn lao chết trộn trong nước và dầu.
(4) Sự di truyền khả năng đáp ứng của cơ thể: cùng một kháng nguyên nhưng các cơ thể khác nhau thì tạo ra các đáp ứng miễn dịch ở mức độ khác nhau. Vì thế mà Landsteiner đã phân biệt hai khái niệm: Tính kháng nguyên
và tính miễn dịch, trong đó: Tính sinh miễn dịch = tính kháng nguyên + khả năng đáp ứng của cơ thể.
4.1.2. Tính đặc hiệu
Tính đặc hiệu của mỗi đáp ứng miễn dịch có được là do mỗi kháng nguyên có một cấu trúc riêng. Tính đặc hiệu của kháng nguyên không phải do toàn bộ phân tử kháng nguyên quyết định, mà do một hoặc nhiều đoạn nhỏ nằm trên phân tử kháng nguyên quyết định. Nhưng đoạn nhỏ này các là quyết định kháng nguyên hay epitop. Epitop có hai chức năng, một là kích thích cơ thể tạo ra đáp ứng miễn dịch đặc hiệu với kháng nguyên đó, và hai là làm vị trí để kháng thể hoặc tế bào lympho mẫn cảm có thể gắn vào một cách đặc hiệu.
Một kháng nguyên protein phức tạp có thể nhiều quyết định kháng nguyên khác nhau, do đó mà nó có thể kích thích tạo ra nhiều loại kháng thể khác nhau cùng một lúc. Tùy theo kháng nguyên có thể phản ứng cùng một lúc với một hay nhiều kháng huyết thanh chứa kháng thể do nó tạo ra mà người ta gọi là kháng nguyên đơn giá hay kháng nguyên đa giá. Trong các phản ứng huyết thanh học chỉ có những kháng nguyên đa giá mới có thể tạo ra mạng lưới kết tủa hoặc ngưng kết.
4.1.3. Phản ứng chéo
Phân tử kháng thể có tính đặc hiệu rất cao, nhưng cũng có trường hợp kháng thể của kháng nguyên A lại tác dụng với kháng nguyên B, ta gọi là phản ứng chéo. Nguyên nhân của phản ứng chéo có thể là do trên hai kháng nguyên này có hai epitop giống nhau hoặc ít nhất là cũng tương tự nhau.
Trong thực nghiệm chúng ta có thể loại trừ được phản ứng chéo bằng phương pháp cho hấp thụ. Ví dụ, ta biết kháng huyết thanh kháng A thường cho phản ứng chéo với kháng nguyên B cho nên khi làm phản ứng tìm kháng nguyên A thì kết tủa dễ sai lạc do tìm nhầm cả
B. Như vậy, trước khi tìm A ta cho ủ kháng nguyên huyết thanh kháng A với kháng nguyên B, nhưng phân tử nào cho phản ứng chéo sẽ tạo phức hợp với B. Sau khi ly tâm loại phức hợp ta sẽ còn kháng huyết thanh A không còn phản ứng chéo với B.
4.1.4. Hapten
Hapten hay bán kháng nguyên là một kháng nguyên không toàn năng, có trọng lượng phân tử thấp, không có tính sinh miễn dịch nhưng có tính đặc hiệu kháng nguyên. Khi hapten được gắn với một chất protein tải thành một phức hợp thì phức hợp này có tính sinh miễn dịch. Nói rò hơn, trong thực nghiệm, nếu ta chỉ đưa hapten vào cơ thể thì
Không có KT
Kháng
Kháng chất
Hình 4.1. Sơ đồ minh họa đáp ứng với hapten
Bản thân hapten không tạo ra đáp ứng, nhưng khi được liên kết in vitro và/hoặc in vivo với cộng hợp protein thì sẽ tạo ra kháng thể phản ứng cả với hapten và với cộng hợp protein đó.
4.2. Một số kháng nguyên quan trọng
4.2.1. Hệ kháng nguyên nhóm máu ABO
Hệ này bao gồm 4 nhóm máu khác nhau: A, B, AB và O. Ký hiệu nhóm máu biểu thị kháng nguyên có mặt trên bề mặt hồng cầu. Cơ thể nhóm máu A thì có kháng nguyên A, nhóm B thì có kháng nguyên B, nhóm AB có kháng nguyên A và B và nhóm O thì không có cả A và B trên hồng cầu. Một đặc điểm khác của cơ thể là người máu A có mang kháng thể chống B trong huyết thanh, người máu B thì có huyết thanh chống A, người AB thì huyết thanh không có kháng thể chống hai kháng nguyên này còn nhóm O thì có cả hai loại kháng thể.
Các kháng nguyên thuộc hệ ABO do một locus gen kiểm soát với ba allel A, B, và O, trong đó A và B trội hơn O. Tính đặc hiệu kháng nguyên nhóm máu trong hệ ABO được quyết định bởi sự có mặt của một số gốc “ose” trong phần polysaccharid. Các kháng nguyên này đều có chung một lòi sphingolipid- polysaccharid. Nếu lòi này được gắn thêm gốc fucose thì xuất hiện chất H. Chất này có trên bề mặt hồng cầu của hầu hết các cơ thể và là nền để suất hiện kháng nguyên A và kháng nguyên B. Nếu tại vị trí galactose cuối cùng của chất H có gắn thêm N-acetyl galactosamin thì xuất hiện kháng nguyên A. Còn nếu gắn thêm một galactose nữa thì xuất hiện kháng nguyên B (Hình 4.2).
Hình 4.2. Sơ đồ cấu tạo kháng nguyên ABO
Các gen A hoặc B mã hóa cho enzym transferase gắn N-acetylgalactosamin hoặc
3
2
OLIGOSACCARIT
galactose vào chất H để tạo nên nhóm máu A
hoặc B. Nếu
không có gen A hoặc B thì máu thuộc nhóm O.
Về mặt di truyền, tham gia vào sự hình thành kháng nguyên nhóm máu có hai hệ gen, đó là hệ Hh đối với chất H và hệ ABO. Hai hệ này hoạt động độc lập với nhau. Đại đa số chúng ta có gen H, nhưng cũng có người không có (cơ thể đồng hợp tử hh). Những người này dù có gen của hệ ABO nhưng trên hồng cầu vẫn không có kháng nguyên A hoặc B nên được ghi nhận là nhóm O nhưng khi truyền máu O thực sự vào người này (tức đưa chất H vào cho người không có H) thì có thể gây tai biến truyền máu. Nhóm đặc biệt này được gọi là nhóm O Bombay.
4.2.2. Hệ kháng nguyên nhóm máu Rh
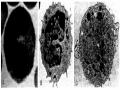
Năm 1930, Landsteinner và Wiener đã phát hiện ra nhóm máu Rh. Họ gọi những người có hồng cầu ngưng kết với huyết thanh thỏ kháng hồng cầu của khỉ Rhessus là người Rh+, số người còn lại là Rh-. Nhưng rồi sau này, người ta đã phát hiện ra rằng, kháng nguyên trong hệ Rh không đơn giản như vậy. Hệ Rh có nhiều kháng nguyên, phần lớn có tính sinh miễn dịch yếu và hay gây phản ứng chéo. Chỉ có kháng nguyên D là có tính sinh miễn dịch mạnh. Khi hồng cầu có kháng nguyên D thì được gọi là Rh+ mà không cần để ý đến các kháng nguyên khác trong hệ này. Kháng thể kháng D không xuất hiện tự nhiễm trong máu như các kháng thể của hệ ABO. Vì kháng nguyên Rh phân bố thưa thớt trên hồng cầu nên thường khó gây ngưng kết hồng cầu khi làm phản ứng xác định nhóm máu Rh, vì thế người ta phải dùng đến thử nghiệm Coombs giám tiếp (Hình 4.3)

Hình 4.3. Phản ứng Coombs gián tiếp
(a). Hình ảnh phản ứng âm tính; (b) Hình ảnh phản ứng dương tính; (c) Cơ chế phản ứng trong trường hợp bất đồng nhóm máu Rh.
4.2.3. Kháng nguyên vi sinh vật
Kháng nguyên vi sinh vật là loại kháng nguyên rất phức tạp: kháng nguyên vỏ, kháng nguyên thân, kháng nguyên lông, kháng nguyên ngoại tế bào, v.v…
Về kháng nguyên vỏ thì kháng nguyên vỏ của phế cầu thường hay được nghiên cứu nhiều nhất. Kháng nguyên này thuộc loại polysaccaride, có tới 80 typ huyết thanh khác nhau.
Về kháng nguyên thân thì ở các vi khuẩn ruột non, lớp ngoài của vách vi khuẩn có một lipopolisaccharide: phần lipid có tính độc, còn phần polysaccharid thì có tính kháng nguyên và đây chính là kháng nguyên thân (còn gọi là kháng nguyên O). Ở Salmonella có đến hơn 60 kháng nguyên O mà cấu trúc có hai phần: phần nhân cơ bản và phần chuổi ngang. Phần nhân giống nhau ở tất cả các Salmonella. Phần chuổi ngang gồm nhưng tiểu đơn vị oligosaccharide sắp xếp lặp đi lặp lại. Chính các chuổi ngang quyết định tính đặc hiệu của mỗi nhóm Salmonella.
Các kháng nguyên lông của Salmonella có bản chất là protein, còn được gọi là kháng thể H, quyết định tính đặc hiệu của mỗi typ Salmonella. Người ta đã phát hiện Salmonella có hơn 1000 typ huyết thanh khác nhau.
Các kháng nguyên ngoại tế bào như độc tố và các enzym cũng có bản chất protein. Một loại kháng nguyên ngoại tế bào của liên cầu là Streptolysin O thường được dùng trong chẩn đoán huyết thanh. Các vi khuẩn trong bạch hầu, uốn ván có ngoại độc tố gây bệnh và các vaccin phòng các bệnh này đây là độc tố đã được giải độc.
Kháng thể virus có thể ở trên bề mặt hạt virus (capsid) hoặc ở bên trong. Tùy theo tính đặc hiệu, có thể phân biệt các kháng nguyên nhóm, các kháng nguyên typ và kháng