- External signals can regulate cytokine receptor expression and thus cytokine response.
- The effect of most cytokines on cells is to create changes in gene expression, causing the appearance of new functions and sometimes causing proliferation of target cells.
Maybe you are interested!
-

 Basic electronic engineering - City College of Construction. HCM Part 1 - 1
Basic electronic engineering - City College of Construction. HCM Part 1 - 1 -
 Business Communication and Negotiation Part 2 - 12
Business Communication and Negotiation Part 2 - 12 -
 Office 2013 Basic Part 2 - 1
Office 2013 Basic Part 2 - 1 -
 Designing and using concept maps in teaching the part "Genetics" contributes to improving the quality of teaching Biology for grades 12 - 27
Designing and using concept maps in teaching the part "Genetics" contributes to improving the quality of teaching Biology for grades 12 - 27 -
 Soft skills - Master Pham Thi Cam Le Part 2 - 1
Soft skills - Master Pham Thi Cam Le Part 2 - 1
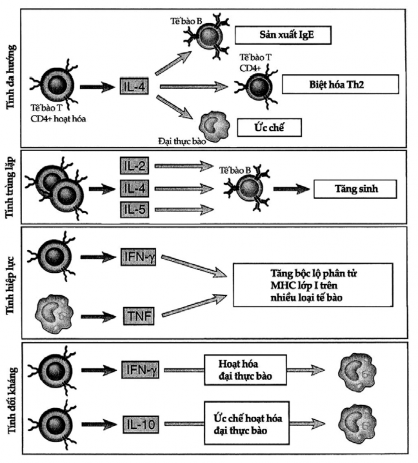
Figure 7.2. Properties of cytokines
Here are examples of the properties of cytokines: pleiotropy: a cytokine can act on many different cells, overlapping effects: many cytokines can have the same effect on the same cell type, synergy: two or more cytokines have a stronger effect than one, and antagonism: one cytokine can inhibit the effect of another.
7.1.2. Functions of cytokines
Cytokines have three main functions:
1. Mediators and regulators of innate immunity: these are cytokines produced by mononuclear phagocytes. The production of these cytokines
These cytokines are stimulated by bacterial products such as lipopolysaccharide (LPS) or viral products such as double-stranded RNA and act as part of the innate immune system. They are also produced by antigen-specific T cells and act as part of the adaptive immune system. They act on endothelial cells and leukocytes to induce an early inflammatory response. NK cells also produce some cytokines.
2. Mediators and regulators of acquired immunity: these are cytokines produced mainly by T lymphocytes to fight foreign antigens. Some cytokines have the main effect of regulating the maturation and differentiation of different lymphocyte populations, so they play an important role in the activation phase of the T cell-dependent response. Some other cytokines originating from T cells are important in the effector phase because they have the function of regulating effector cell types such as phagocytes, neutrophils, eosinophils, etc.
3. As hematopoietic stimulants: these cytokines are produced by stromal cells, leukocytes, and some other cells of the bone marrow. They can stimulate the growth and differentiation of immature leukocytes.
In summary, cytokines of the innate and adaptive immune systems are often produced by different cell types and act on different target cell types. However, this distinction is not absolute because a cytokine can be produced in both the innate and the adaptive response, and different cytokines can produce some of the same effects.
7.1.3. Cytokine receptors and signaling
All cytokine receptors have one or more transmembrane proteins and an extracellular domain that binds to the cytokine molecule to initiate a signal cascade into the cell. Cytokine receptors are classified based on the structural similarity of their extracellular cytokine-binding domains. There are five families of cytokine receptors (Figure 7.3A).
1. Type I cytokine receptors: also known as hematopoietic receptors, contain one or more copies of a domain with two pairs of fixed cysteine residues and a juxtamembrane sequence tryptophan-serine-X-tryptophan-serine (WSXWS), where X is any amino acid. This type of receptor recognizes cytokines that are coiled into four α- helices . These receptors have specific ligand-binding sequences and one or more signaling sequences that are structurally similar when recognizing different cytokines (Figure 7.3B).
2. Type II cytokine receptor: This type is similar to type I in that it has two extracellular domains that carry cysteine-fixing molecules, but type II lacks the WSXWS juxtamembrane sequence. This type of receptor has a ligand-binding polypeptide chain and a signal transduction sequence.
3. Some cytokine receptors have extracellular Ig domains and are classified into the Ig superfamily. This group of receptors binds to different cytokines and transmits signals by different mechanisms.
4. The TNF receptor belongs to a family of receptors (including many non-cytokine receptors) that have extracellular domains containing multiple cysteine-fixing molecules. Upon binding to ligand, these receptors activate a number of intracellular proteins to induce apoptosis or stimulate gene expression, or both.
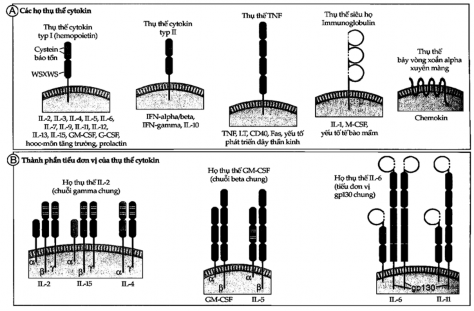
Figure 7.3A and B. Structure of cytokine receptors
A. Cytokine receptors are classified into several families based on their extracellular matrix.
B. Cytokine receptors are composed of cytokine-specific ligand-binding chains (usually α chains ) noncovalently linked to signaling subunits whose structures may be identical.
5. Heptameric receptors are also called snake receptors because their transmembrane domains are shaped like snakes running across the membrane. In the immune system, these receptors mediate rapid and transient responses to a family of cytokines called chemokines.
7.2. Cytokines mediate and regulate innate immunity
7.2.1. TNF (tumor necrosis factor)
TNF is a major mediator of acute inflammatory responses against gram-negative bacteria and some other microorganisms. TNF is also responsible for many of the systemic complications of severe infections. TNF was first discovered in the serum of animals treated with bacterial endotoxin (LPS) and has been shown to induce necrosis of tumors in vivo when present in large amounts.
7.2.1.1. Origin, receptor
The main source of TNF production is mononuclear phagocytes, however, some other cells such as T lymphocytes, NK cells, and mast cells can also secrete this substance. The strongest stimulus for macrophages to produce TNF is LPS. In addition, large amounts of this cytokine are also produced by gram-negative bacteria. Interferon produced by T and NK cells also has the effect of amplifying TNF biosynthesis of macrophages stimulated by LPS.
There are two types of TNF receptors, the 55 kD type called THF-RI and the 75 kD type called TNF-RII. These receptors are present on most cell types in the body.
7.2.1.2. Biological activity
- The main physiological function of TNF is to stimulate the concentration of neutrophils and monocytes to the site of infection and activate these cells to destroy bacteria (Figure 7.4).
TNF exposes adhesion molecules on endothelial cells that cause leukocytes, neutrophils, and monocytes to adhere to the endothelial cell. Two important adhesion molecules here are selectins and ligands for leukocyte integrins.
TNF stimulates endothelial cells and macrophages to secrete chemokines that enhance the affinity of leukocyte integrins for their ligands and induce leukocyte aggregation. TNF also stimulates mononuclear phagocytes to secrete IL-1, which has effects very similar to TNF.
In addition to its role in inflammation, TNF also initiates programmed cell death in some cells.
- In severe infections, TNF is produced in large amounts and causes systemic clinical symptoms and pathological lesions. If the stimulation of TNF production is strong enough, it can cause overproduction of TNF and this amount will spill into the blood to cause effects far from the site of infection like a hormone. Systemic effects of TNF include:
+ TNF acts on the hypothalamus to cause fever, so it is called an endogenous pyrogen (to distinguish it from LPS, which is an exogenous pyrogen). Fever caused by TNF (and also IL-1) is mediated by prostaglandin biosynthesis. Therefore, anti-prostaglandin agents can reduce fever caused by TNF and IL-1.
+ TNF acts on liver cells to increase serum protein synthesis. Plasma proteins produced by the liver under the influence of TNF, IL-1 and IL-6 create an acute phase response of the inflammatory reaction.
+ Prolonged TNF production causes muscle and fat cell wasting and eventually leads to cachexia.
+ When a large amount of TNF is produced, the ability of the heart muscle and vascular smooth muscle to contract is inhibited, causing hypotension and sometimes shock.
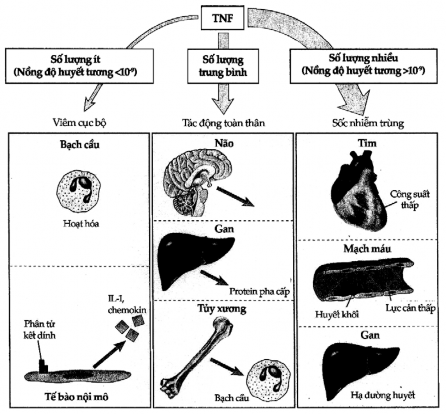
Figure 7-4. Biological activities of TNF
At low concentrations, TNF acts on leukocytes and endothelium to initiate an inflammatory response. At intermediate concentrations, TNF mediates the systemic effects of the inflammatory response. And at high concentrations, TNF causes the pathological abnormalities of septic shock.
+ TNF causes intravascular thrombosis due to endothelial cell loss of properties
normal anticoagulant. TNF stimulates endothelial cells to express tissue factor, a potent coagulation promoter. Its ability to induce tumor necrosis, as its name suggests, results from thrombosis in tumor blood vessels.
+ Large amounts of TNF circulating in the blood can also cause metabolic disorders, specifically hypoglycemia due to increased sugar consumption but the liver cannot compensate.
A serious complication of gram-negative sepsis is septic shock (also known as endotoxic shock). The clinical picture of shock includes cardiovascular collapse, disseminated intravascular coagulation, and metabolic disorders. This syndrome is caused by the overproduction of TNF and other cytokines such as IL-12, INF- , and IL-1 under the influence of bacterial LPS. Measurement of serum TNF levels can predict the clinical picture of this shock. TNF antagonists can prevent death in experimental models but not in clinical practice, and the reason is probably that other cytokines besides TNF have the same effect as TNF and are equally potent.
7.2.2. Interleukin-1 (IL-1)
The primary function of IL-1, like TNF, is to mediate the host's inflammatory response to infection and other inflammatory stimuli. IL-1 functions together with TNF in the innate immune system.
7.2.2.1. Origin, receptor
The primary source of IL-1, like TNF, is activated mononuclear phagocytes. But unlike TNF, IL-1 is also produced by a number of other cells such as neutrophils, epithelial cells, and endothelial cells.
There are two forms of IL-1 called IL-1 and IL-1 , both of which bind to the same cell receptor and have similar biological activity. Both are produced as a 33 kD precursor and a 17 kD cleaved fragment. The active form of IL-1 is the cleaved fragment while the active form of IL-1 is both the precursor and the cleaved fragment. The majority of IL-1 found in the circulation is IL-1 .
There are two membrane receptors for IL-1 called type I and type II receptors. Both belong to the Ig superfamily. Type I receptors are found on most cells, while type II receptors are found primarily on B cells, but can also stimulate production on some other cells.
7.2.2.2. Biological activity
The biological activity of IL-1 is similar to that of TNF and depends on the amount produced.
When secreted at low concentrations, IL-1 acts as a mediator of local inflammation. It acts on endothelial cells to increase the expression of leukocyte adhesion surface molecules such as integrin ligands.
When secreted in large amounts, IL-1 enters the bloodstream and acts as a hormone. The systemic effects of IL-1 are similar to those of TNF: fever, stimulation of the liver to produce acute-phase plasma proteins, and cachexia.
The similarities in the actions of IL-1 and TNF are striking. However, they also differ: IL-1 does not mediate programmed cell death (apoptosis) even at high concentrations and does not itself cause septic shock.
Mononuclear phagocytes produce a natural inhibitor of IL-1. This substance shares the same receptor with IL-1 and acts as a competitive inhibitor of IL-1, hence the name IL-1 receptor antagonist (IL-1ra). IL-1ra may be an endogenous regulator of IL-1.
7.2.3. Chemokines
Chemokines are a large family of cytokines that stimulate leukocyte migration and regulate their movement from the blood to tissues. The word chemokine is short for chemotactic cytokine. Some chemokines can be produced by many different cell types.
in response to inflammatory stimuli and attract leukocytes to the site of inflammation. Several other chemokines are produced by various tissues even in the absence of inflammation and also attract leukocytes (mainly lymphocytes) to the tissue.
7.2.3.1. Structure, origin, receptors
All chemokines are polypeptides with a molecular weight of 8-12 kD and contain two disulfide bridges inside. About 50 chemokines have been identified and more may be discovered in the future. Chemokines are divided into several families based on the number of N-terminal cysteine molecules. The two main families are the CC chemokine family, which has terminal cysteine molecules located next to each other, and the CXC family, which has terminal cysteine molecules separated by one amino acid. In inflammatory reactions, CXC chemokines act mainly on neutrophils, while CC acts on monocytes, lymphocytes, and eosinophils. Chemokines also have a C family, which has only one cysteine molecule, or a CX3C family, which has two cysteine molecules separated by three amino acids.
In origin, chemokines involved in inflammatory responses are produced by leukocytes in response to external stimuli. Meanwhile, chemokines that regulate cell traffic through tissues are produced by different cells within these tissues.
Eleven chemokine CC receptors have been identified, named CCR1 to CCR11, and six CXC receptors, named CXC1 to CXC6, and this is not all. Chemokine receptors are expressed on leukocytes, especially on T cells, where many chemokine receptors are found. All chemokine receptors share a common characteristic structure consisting of domains with a seven-helix structure across the membrane.
There are several chemokine receptors such as CCR5 and CXCR4 that act as co-receptors for HIV. Some activated T lymphocytes can secrete chemokines that bind to CCR5 and compete with the virus, thereby blocking HIV infection.
7.2.3.2. Biological activity
Chemokines were discovered for their chemotactic function but they actually have many other important functions in the immune system as well as other systems.
- Chemokines attract host cells to the site of infection. Chemokines present on endothelial cells act on passing leukocytes, causing leukocyte integrins to increase their binding affinity for their ligands. This is important for retaining leukocytes in the endothelium of capillaries in the damaged area. In addition, chemokines also stimulate leukocyte migration to the site of injury based on the concentration gradient of chemokines at the site of injury and elsewhere. Different chemokines stimulate different cells, thereby controlling the composition of cells at the site of inflammation. For example,
The chemokine IL-8 mobilizes mainly neutrophils, while the chemokine eotaxin concentrates eosinophils.
- Chemokines regulate the trafficking of lymphocytes and other leukocytes in peripheral lymphoid tissues. This is a new discovery in immunology. Chemokines have the ability to promote the migration of activated effector cells and memory T cells to non-lymphoid tissues including skin and mucous membranes. The selection of different cells to different tissues depends on the number and cellular receptors of each chemokine.
7.2.4. Interferon (IFN) type I
Type I interferons are mediators of the early response of innate immunity against viral infections.
7.2.4.1. Structure, origin and receptors
Type I IFNs consist of two groups of proteins, IFN- and IFN- . IFN- is a family of 20 structurally related polypeptides, each encoded by a separate gene. IFN- is produced primarily by mononuclear phagocytes and is sometimes called leukocyte interferon. IFN- is a protein produced by many cell types, such as fibroblasts, and is therefore sometimes called fibroblast interferon. The most effective stimulus for interferon synthesis is viral infection, especially double-stranded RNA viruses produced by viruses during their replication in host cells. In vitro, type I IFN production is stimulated by artificial double-stranded RNAs that have signals very similar to those of viral infections. Antigen-activated T cells also stimulate mononuclear phagocytes to synthesize type I IFN.
Although IFN- α and IFN- β are structurally different, they bind to the same receptor and induce similar biological responses.
7.2.4.2. Biological activity
The activity of type I IFN is to protect the body against viral infections and to promote cellular immune responses against intracellular microorganisms. (Figure 7.5).
- Type I IFN inhibits viral replication. IFN stimulates cells to produce many enzymes such as 2'5' oligoadenylate synthetase, which prevents the replication of DNA or RNA viruses and inhibits their replication. The antiviral effect of type I IFN is both autocrine, meaning it inhibits viral replication within itself, and paracrine, meaning it protects neighboring uninfected cells.
- Type I IFN has the effect of increasing the expression of MHC class I molecules. CD8+ T cells have the ability to recognize foreign antigens associated with MHC class I,